செல் கொள்கை:
1833-ஆம் ஆண்டு ஜெர்மனி தாவரவியலார் மாத்தியோஸ் ஷிலீடன், ஜெர்மனி விலங்கியலார் தியோடர் ஷிவான் இருவரும் சேர்ந்து அனைத்துத் தாவரங்களும் விலங்குகளும் செல்களாலானவை என்றும், இச்செல்கள்தான் உயிரினங்களின் அடிப்படை அலகாகத் திகழ்கின்றன என்றும் கூறினர்.
அனைத்து உயிரினங்களும் செல்களால் ஆனவை.
ஏற்கனவே உள்ள செல்களிலிருந்து புதிய செல்கள் தோன்றுகின்றன.
செல் மரபியல் தகவல்களைக் கொண்டுள்ளது. இவை பெற்றோரிடமிருந்து சந்ததிகளுக்குக் கடத்தப்படுகிறது.
அனைத்து வளர் சிதை மாற்ற வினைகளும் செல்லுக்குள்ளே நடைபெறுகிறது.
செல் கொள்கையின் விதிவிலக்கு:
வைரஸ்கள் உயிரியல் வல்லுநர்களுக்கு ஒரு புதிராகவே இருந்தன. வைரஸ்கள், வைராய்டுகள், பிரியான்கள் ஆகியவை செல்கொள்கைக்கு ஒரு விதி விலக்காகும். செல்லின் முக்கியப் பகுதியான புரோட்டோபிளாசம் அவைகளுக்கு இல்லை. மேலும் இவை செல்லுக்குள் வாழும் கட்டாய ஒட்டுண்ணிகளாக இருக்கின்றன.
செல்லின் வகைகள்:
செல்லின் ஒழுங்கமைவுமற்றும்உட்கரு பண்பினைக் கொண்டு உயிரினங்கள் கீழ்க்கண்டவாறு வகைப்படுத்தப்படுகின்றன.
புரோகேரியோட்டுகள்
தொன்மையான உட்கரு கொண்ட உயிரிகள் புரோகேரியோட்டுகள் எனப்படும் (Pro தொன்மையான Karyon-உட்கரு). புரோகேரியாட்டுகளில் “நியூகிளியாய்டு” பகுதியில் ஹிஸ்டோன் புரதம் அற்ற DNA உட்கரு சவ்வு அற்று காணப்படுகிறது. ஆகையினால் இது உண்மையான உட்கரு அன்று. எடுத்துக்காட்டு: பாக்டீரியங்கள், நீலப்பசும்பாசிகள், மைக்கோபிளாஸ்மா, ரிக்கெட்சியே மற்றும் ஸ்பைரேகிட். மேலும் இதன் உட்கரு பொருட்கள் தொன்மையானவை.
மீசோகேரியோட்டுகள்
டாட்ஜ் என்னும் அறிவியலாளர் மற்றும் அவரது சக ஆராய்ச்சியாளர்கள் மூன்றாவது வகை உயிரினங்களை மீசோகேரியோட்டுகள் என்று அழைத்தனர். புரோகேரியோட்டின் சில பண்புகளையும், யூகேரியோட்டின் சில பண்புகளையும் இந்த உயிரிகள் பெற்றுள்ளன. இந்த மீசோகேரியோட்டுகள், புரோகேரியோட்டுகள் மற்றும் யூகேரியோட்டுகளுக்கும் இடைப்பட்டவைகளாக காணப்படுகின்றன. இவற்றில் நன்கு உருவாகிய உட்கரு சவ்வால் சூழப்பட்டுள்ளது. இதன் DNA குரோமோசோம்களாகவும், ஹிஸ்டோன் புரதமற்றும் காணப்படுகிறது. இவைகள் புரோகேரியோட்டுகளைப் போல நேர்முகப்பிரிவு பகுப்பைக் கொண்டுள்ளன. எடுத்துக்காட்டு. நாக்டியூலியா என்ற புரோடோசோவா மற்றும் ஜிம்னோடினியம், பெரிடினியம் போன்ற தாவர மிதவை உயிரிகள் மற்றும் டைனோபிளஜெல்லேட்டுகள்.
யூகேரியோட்டுகள்
உண்மையான உட்கருவைக் கொண்ட உயிரிகளுக்கு யூகேரியோட்டுகள் என்று பெயர். (Eu-உண்மை, Karyon-உட்கரு). இதன் குரோமோசோம்களில் உள்ள DNA வானது ஹிஸ்ட்டோன் புரதங்களால் ஆனவை. சவ்வு சூழ்ந்த பல செல் நுண்ணுறுப்புக்களைக் கொண்டவை. உள்ளுறை கூட்டுயிர் வாழ்க்கை முறை மூலம் இந்த உள் உறுப்புக்கள் தோன்றி ஒரு செல் மற்றொரு செல்லினுள் இருப்பதுபோல வாழ்கின்றன. மைட்டோகாண்ட்ரியங்களும், பசுங்கணிகங்களும் இந்தக் கூட்டுயிர் வாழ்க்கை கோட்பாட்டை உறுதிப்படுத்துவதாக உள்ளன.
செல் அமைவு வகைகளின் ஒப்புமைகள்:
பண்புகள் | புரோகேரியோட்டுகள் | மீசோகேரியோட்டுகள் | யூகேரியோட்டுகள் |
செல்லின் அளவு | ~1-5 µm | ~5-10µm | ~10-100µm |
உட்கருவின் பண்பு | நியூக்ளியாய்டு, உண்மையான உட்கரு அற்றது. | சவ்வுடன் கூடிய உட்கரு காணப்படுகிறது. | சவ்வுடன் கூடிய உட்கரு காணப்படுகிறது. |
DNA | பொதுவாக வட்ட வடிவம், ஹிஸ்டோன் புரதம் அற்றவை. | பொதுவாக நீள் வடிவம், ஹிஸ்டோன் புரதம் அற்றவை. | பொதுவாக நீள் வடிவம், ஹிஸ்டோன் புரதம் கொண்டவை. |
RNA உற்பத்தி, புரதச்சேர்க்கை | சைட்டோபிளாசத்தில் நடைபெறுகிறது. | யூகேரியோட்டுகளை ஒத்துள்ளன. | RNA உட்கருவினுள் உருவாகின்றது புரதச்சேர்கை சைட்டோபிளாசத்தினுள் நடைபெறுகிறது. |
ரைபோசோம்கள் | 50S + 30S | 60S + 40S | 60S + 40S |
நுண்ணுறுப்புகள் | இல்லை. | உள்ளன. | பல காணப்படுகிறது. |
செல் இடப்பெயர்ச்சி | கசையிழை. | இழைந்து நழுவுதல் மற்றும் கசையிழை. | கசையிழை மற்றும் குறுஇழை |
அமைவு முறை | பொதுவாக ஒற்றைச் செல். | ஒற்றைச் செல் மற்றும் கூட்டமைவு. | ஒற்றைச் செல், கூட்டமைவு மற்றும் பல செல்களைக் கொண்டது. |
செல் பகுப்பு | இருபிளவுறுதல் முறை. | இருபிளவுறுதல் முறை. | மைட்டாசிஸ், மியாசிஸ். |
எடுத்துக்காட்டுகள் | பாக்டீரியா மற்றும் ஆர்க்கியா. | டைனோபிளஜெல்லேட்டுகள், புரோடோசோவா. | பூஞ்சை, தாவரங்கள் மற்றும் விலங்குகள். |
தாவர மற்றும் விலங்கு செல்:
யூகேரியோட்டிக் செல்கள் நன்கு தெளிவான அமைப்பைக் கொண்டுள்ளன. இவை உயிரினங்களில் பல வேறுபாடுகளுடன் காணப்படுகின்றன. எடுத்துக்காட்டாகத் தாவர மற்றும் விலங்கு செல்கள்.
விலங்கு செல்:
விலங்கு செல்லானது செல் சவ்வு அல்லது பிளாஸ்மா சவ்வினால் சூழப்பட்டிருக்கிறது. இந்தச் சவ்வினுள் புரோட்டோபிளாசம் என்னும் ஜெலாடினஸ் மாட்டிரிக்ஸ் காணப்படுகிறது. இதில் உட்கரு மற்றும் எண்டோபிளாச வலை, மைட்டோகாண்டிரியா, கோல்கை உடலம், சென்ட்ரியோல்கள், லைசோசோம்கள், ரைபோசோம்கள் மற்றும் செல் சட்டகம் போன்ற பிற உறுப்புகளும் உள்ளன.
தாவர செல்:
தாவரச் செல்கள் வழக்கமான, தெளிவான செல் சுவரையும், ஒரு பெரிய மைய வாக்குவோலையும் மற்றும் கணிகங்களையும் பெற்றுள்ளன. மேலும் விலங்கு செல்களில் உள்ளது போல் பல செல் நுண்ணுறுப்புகளும் காணப்படுகின்றன.
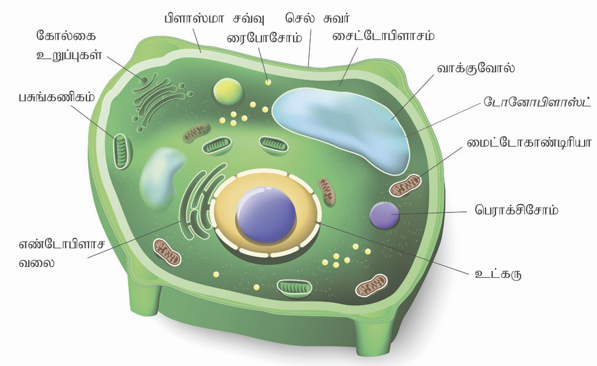
செல்லின் நுண்ணுறுப்புகள் மற்றும் அதன் பணிகள்
வ. எண் | செல்லின் பாகம் | முக்கியப் பணிகள் | சிறப்புப் பெயர் |
1 | செல் சுவர் | செல்லைப் பாதுகாக்கிறது. செல்லிற்கு உறுதி மற்றும் வலிமையைத் தருகிறது. | தாங்குபவர் (அல்லது) பாதுகாப்பவர். |
2 | செல் சவ்வு | செல்லிற்குப் பாதுகாப்பு தருகிறது. செல்லின் போக்குவரத்திற்கு உதவுகிறது | செல்லின் கதவு |
3 | சைட்டோபிளாசம் | நீர் அல்லது ஜெல்லி போன்ற, செல்லில் உள்ள நகரும் பொருள் | செல்லின் நகரும் பகுதி |
4 | மைட்டோ காண்டிரியா | செல்லிற்குத் தேவையான அதிக சக்தியை உருவாக்கித் தருகிறது | செல்லின் ஆற்றல் மையம் |
5 | பசுங்கணிகம் | இதில் பச்சையம் என்ற நிறமி உள்ளது. இது சூரிய ஒளியை ஈர்த்து ஒளிச் சேர்க்கையின் மூலம் உணவு தயாரிக்க உதவுகிறது. | செல்லின் உணவுத் தொழிற்சாலை |
6 | நுண்குமிழ்கள் | இது உணவு, நீர் மற்றும் வேதிப் பொருள்களைச் சேமிக்கிறது. | சேமிப்புக் கிடங்கு |
7 | உட்கரு (நியூக்ளியஸ்) | செல்லின் மூளையாகச் செயல்படுகிறது. செல்லின் அனைத்துச் செயல்களையும் ஒருங்கிணைத்துக் கட்டுப்படுத்துகிறது. | செல்லின் கட்டுப்பாட்டு மையம் |
8 | உட்கரு உறை (நியூக்ளியஸ் உறை) | நியூக்ளியஸைச் சுற்றி அமைத்து, அதைப் பாதுகாக்கிறது நியூக்ளியஸின் உள்ளேயும் வெளியேயும் பொருள்களை அனுப்புகிறது. | உட்கரு வாயில் (அல்லது) உட்கரு கதவு |
தாவர மற்றும் விலங்கு செல்களுக்கு இடையிலான வேறுபாடு:
வ.எண் | தாவர செல் | விலங்கு செல் |
1 | பொதுவாக அவை விலங்கு செல்களை விட பெரியவை | பொதுவாக தாவர செல்களை விட சிறியது |
2 | பிளாஸ்மா மென்படலத்துடன் கூடுதலாக செல் சுவர் உள்ளது மற்றும் நடுத்தர லேமல்லே, முதன்மை மற்றும் இரண்டாம் நிலை சுவர்கள் உள்ளன | செல் சுவர் இல்லை |
3 | பிளாஸ்மோடெஸ்மாட்டா உள்ளது | பிளாஸ்மோடெஸ்மாட்டா இல்லை |
4 | குளோரோபிளாஸ்ட் உள்ளது | குளோரோபிளாஸ்ட் இல்லை |
5 | வெற்றிடமானது பெரியது மற்றும் நிரந்தரமானது | வெற்றிடமானது சிறியது மற்றும் தற்காலிகமானது |
6 | வெற்றிடத்தைச் சுற்றி டோனோபிளாஸ்ட் உள்ளது | டோனோபிளாஸ்ட் இல்லை |
7 | கீழ் தாவரங்களின் அசையும் செல்களைத் தவிர சென்ட்ரியோல்கள் இல்லை | சென்ட்ரியோல்கள் உள்ளன |
8 | அணுக்கரு செல்லின் சுற்றளவில் உள்ளது | செல்லின் மையத்தில் உள்ள கரு |
9 | லைசோசோம்கள் அரிதானவை | லைசோசோம்கள் உள்ளன |
10 | சேமிப்பு பொருள் ஸ்டார்ச் தானியங்கள் | சேமிப்பகப் பொருள் கிளைகோஜன் துகள்கள் |
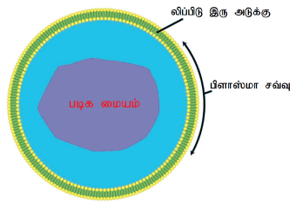
புரோட்டோபிளாசம்:
புரோட்டோபிளாசம் செல்லின் உயிருள்ள பொருள். இது பிளாஸ்மா சவ்வினால் சூழ்ந்து காணப்படுகிறது. இது ஒரு நிறமற்ற பொருளாகும். மேலும்இதுசெல்முழுவதும் பரவி, சைட்டோபிளாசம், உட்கரு மற்றும் பல உள்ளுறுப்புக்களைக் கொண்டுள்ளன. புரோட்டோபிளாசம் சிறிய துகள்களான அயனிகள், அமினோ அமிலங்கள், ஒற்றைச் சர்க்கரைகள் மற்றும் நீரையும், பெரும மூலக்கூறுகளான நியூக்ளிக் அமிலங்கள், புரதங்கள், லிப்பிடுகள் மற்றும் பல்கூட்டுச் சர்க்கரைகள் போன்ற கூட்டுப் பொருள்களை உள்ளடக்கியது. இவை நிறமற்ற தோற்றத்துடன் ஜெல்லி போன்ற மீள்பாகு நிலை கொண்ட துகள்களால் ஆனது. இவை அதிக எண்ணிக்கையிலான வாக்குவோல்கள் கொண்டுள்ளதால் நுரை போன்று காணப்படுகிறது. இது வெப்பம், மின் அதிர்ச்சி, வேதிப்பொருள் ஆகியவற்றின் தூண்டுதலுக்கு ஏற்பத் துலங்கலாகச் செயல்படுகிறது.
செல் சுவர்:
செல்சுவர் செல்லின் வெளிப்பகுதியில் காணப்படும் பாதுகாப்பு அடுக்கு ஆகும். இது பாக்டீரியா, பூஞ்சை, தாவரங்கள் ஆகியவற்றில் காணப்படுகிறது. ஆனால் விலங்கு செல்லில் காணப்படுவதில்லை. இதனை முதன்முதலில் இராபர்ட் ஹுக் என்பவர் உற்று நோக்கினார். இது தொடர்ந்து வளர் ந்து வரும் பகுதியாகும். இது உயிரினங்களில் பல்வேறு கூட்டுப்பொருள்களைக் கொண்டிருக்கிறது. பாக்டீரியங்களில் செல்சுவர் பெப்டிடோகிளைக்கானால் ஆனது. பூஞ்சைகளில் இது கைட்டின் மற்றும் பூஞ்சை செல்லுலோஸினால் ஆனது. ஆல்காக்க ளில்செல்லுலோஸ், கேலக்டான்ஸ், மன்னான்ஸ் ஆகியவற்றால் ஆனது. தாவரச் செல்சுவர்கள் செல்லுலோஸ், ஹெமி செல்லுலோஸ், பெக்டின், லிக்னின், கியூட்டின், சூபரின் மற்றும் சிலிக்காவால் ஆனது.
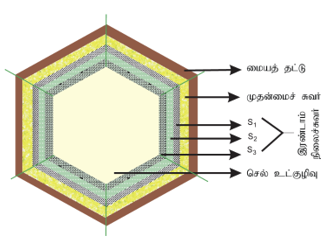
பாகங்கள்:
முதன்மைச்சுவர்:
மையஅடுக்குக்குஉட்புறமாகத் தோற்றுவிக்கப்படும் முதல் அடுக்கு செல்சுவரின் முதன்மைச்சுவராகும். முதன்மைச் செல் சுவரிலுள்ள ஜெல் போன்ற தளப்பொருளில் செல்லுலோஸ் நுண் இழைகள் மிகத் தொய்வாக வலைப்பின்னலைப் போன்று காணப்படுகின்றது. இது மெல்லிய, நீட்சி அடையும் தன்மை உடையது. பெரும்பாலான தாவரங்களில் இந்த நுண் இழைகள் செல்லுலோஸினால் ஆனது. மேலும் சுவரின் வடிவம் மற்றும் தடிமனுக்குத் தக்கவாறு இந்த நுண் இழைகள் பல்வேறு திசையில் அமைந்துள்ளன. முதன்மைச் சுவரின் நுண்பொருள் பெரும்பாலும் ஹெமி செல்லுலோஸ், பெக்டின், கிளைக்கோபுரதம் மற்றும் நீர் நிரப்புப்பொருளா க உள்ளது. ஹெமிசெல்லுலோஸ் தளப் பொருளுடன் நுண் இழைகளைப் பிணைக்கிறது. கிளைக்கோ புரதங்கள் நுண் இழைகளின் அமைவைத் தீர்மானிக்கிறது. பாரங்கைமா செல்கள் மற்றும் ஆக்குத்திசுக்கள் ஆகியவை முதன்மைச் சுவரை மட்டுமே பெற்றுள்ளன.
இரண்டாம் நிலைச்சுவர்:
செல் முதிர்ச்சி அடைந்தவுடன் இரண்டாம் நிலை செல்சுவர் உருவாக்கப்ப டுகிறது. இது செல் வடிவத்தைத் தீர்மானிப்பதில் முக்கியப் பங்கு வகிக்கிறது. இது தடிமனானது, நீட்சி அடையும் தன்மையற்றது. இவை செல்லுலோஸ் மற்றும் லிக்னினால் ஆனது. இரண்டாம் நிலைச்சுவர் மேலும் மூன்று துணை அடுக்குகளாகப் பிரிகின்றது. இவை முறையே S1, S2 மற்றும் S3 ஆகும். இரண்டாம் நிலைச்சுவரின் நுண் இழைகள் பலதிசைகளில் மென்தகடுகள் போன்று மிக நெருக்கமாக அமைந்து செல்சுவருக்கு வலிமையை அதிகரிக்கின்றது.
மையத்தட்டு:
இது சைட்டோபிளாச பகுப்பின்போது கால்சியம் மற்றும் மெக்னீசியம் பெக்டேட்டுக்கள் படிந்து உருவான வெளிப்புற அடுக்காகும். இது அருகருகே உள்ள இரண்டு செல்களுக்கிடையே மெல்லிய வடிவமில்லாத சிமெண்ட் போன்ற அடுக்காகும். இது ஒளிமுறிவுத்தன்மை (Isotropic) கொண்டது.
பிளாஸ்மோடெஸ்மேட்டா மற்றும் குழிகள்:
செல்சுவர் முழுமையாக இல்லாமல் ஆங்காங்கே குறுகிய துளைகள் உள்ளன. இதற்குப் பிளாஸ்மோடெஸ்மேட்டா என்று பெயர். இது அருகருகே உள்ள செல்களின் புரோட்டோபிளாசத்திற்கு இடையே அமைந்து, இதன் வழியே பல பொருட்கள் செல்வதற்கு ஏதுவாகிறது. செல்சுவரின் சில பகுதிகளில் இரண்டாம்நிலை சுவரடுக்குகள் சீரற்றதாகவும் ஆனால் முதன்மைச் சுவரும், மையத்தட்டும் சீரானதாகவும் காணப்படுகின்றன. இந்தச் சீரற்ற பகுதிக்குக் குழிகள் (Pits) என்று பெயர். அருகருகே உள்ள செல்களின் குழிகள் ஒன்றுக்கொன்று எதிரெதிராக உள்ளன. ஒவ்வொரு குழிக்கும் குழி அறை மற்றும் குழிச் சவ்வு உள்ளன. குழிச்சவ்வில் பல நுண்ணிய துளைகள் உள்ளதால் இவற்றின் வழியே பொருள்கள் எளிதில் ஊடுருவிச் செல்லும். குழிகள் எளிய குழிகள் (Simple pits) மற்றும் வரையுற்ற குழிகள் என இருவகைப்படும்.
செல்சுவரின் பணிகள்:
- செல்லுக்கு ஒரு குறிப்பிட்ட வடிவத்தையும், வலுவையும் அளிக்கிறது.
- பல மூலக்கூறுகள் செல்லினுள் நுழைவதைத் தடுப்புசுவர் (Barrier) போன்றுதடை செய்கின்றன.
- செல்லுக்குள்ளே உள்ள புரோட்டோபிளாசத்தை சேதமடையாமல் பாதுகாக்கிறது.
- ஆஸ்மாட்டிக் அழுத்தம் காரணமாக அதிக நீர் செல்லுக்குள்ளே சென்று அதனால் செல் வெடித்துவிடுவதைத் தடுக்கிறது.
- செல்லைப் பாதுக்காக்கும் முக்கியப் பணியையும் மேற்கொள்கிறது.
செல் சவ்வு:
செல் சவ்வானது செல் பரப்பு அல்லது பிளாஸ்மாச் சவ்வு எனவும் அழைக்கப்படுகிறது. இது ஒரு மெல்லிய அமைப்பாக இருந்து சைட்டோசால் என்ற சைட்டோபிளாச உட்பொருளைக் கட்டுக்குள் வைக்க உதவுகிறது. இது 10 nm அளவிற்கும் குறைவான மெல்லிய சவ்வாகும்.
பாய்ம திட்டு மாதிரி (Fluid Mosaic Model):
ஜோனத்தான் சிங்கர் மற்றும் கார்த் நிக்கோல்சன் ஆகியோர் பாய்ம திட்டு மாதிரியை முன்மொழிந்தனர்.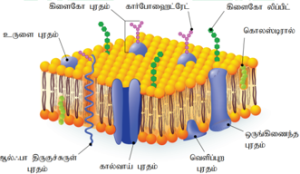
கார்போஹைட்ரேட்டை மிகக் குறைவாகவும், மேலும் லிப்பிடுகள் மற்றும் புரதங்களையும் இது பெற்றுள்ளது. இதில் உள்ள லிப்பிடு சவ்வு பாஸ்போலிப்பிடுகளால் ஆக்கப்பட்டுள்ளது. ஒவ்வொரு பாஸ்போலிப்பிடு மூலக்கூறும் நீர் வெறுக்கும் தன்மை பெற்ற வால் பகுதியையும், நீர் விரும்பும் தலைப்பகுதியையும் கொண்டுள்ளது. நீர் வெறுக்கும் தன்மை பெற்ற வால் பகுதி நீரை வெறுக்கிறது. நீர் விரும்பும் பகுதியானது நீரை ஈர்க்கிறது. இந்த லிப்பிடு மூலக்கூறுகள் சவ் வின் இருவரிசை அடுக்குகளில் அமைந்துள்ளன. இவற்றிற்கு இடையே கோளப் புரத மூலக்கூறுகள் செருகப்பட்டுள்ளது. இப்புரதங்கள் இடைச்செருகு புரதங்கள் எனப்படுகின்றன.
ஒரு சில புரதங்கள் லிப்பிடு அடுக்கின் பரப்பில் காணப்படுகின்றன. இவை வெளியமை புரதங்கள் எனப்படுகின்றன. சவ்வின் வழியே நொ திகள், எதிர் உயிர் பொ ருட்கள் மற்றும் செல்லுக்குத் தேவை யான மூலக்கூறுகள் அனைத்தையும் ஊடு கடத்த இப்புரதங்கள் உதவுகின்றன. சவ்வில் காணப்படும் கார்போஹைட்ரேட்டுகள் குட்டையான கார்பன் சங்கிலியைப் பெற்ற பாலிசாக்ரைடுகளாக உள்ளன. இவை கிளைக்கோபுரதங்கள் அல்லது கிளைக்கோலிப்பிடுகளுடன் பிணைந்து கிளைக்கோகேலிக்ஸ் என உருவாகிறது.
சவ்வில் உள்ள லிப்பிடு பொருட்கள் சவ் வின் ஒரு புறத்திலிருந்து மறுபுறத்திற்குச் செங்குத்து வாக்கில் இடப்பெயரும் தன்மைக்கு அங்கும் இங்குமாக நிகழும் இடப்பெயர்வு (Flip Flop movements) என்று பெயர். பக்கவாட்டில் பரவும்லிப்பிடுமூலக்கூறுகளை விட இந்த இடப்பெயர்வு மிகவும் மந்தமாக நடைபெறுகிறது. பாஸ்போலிப்பிடுகளில் துருவத்தன்மை கொண்ட, மிகச் சிறிய தலைப்பகுதி இருப்பதால்இவை அங்கும்இங்கும்இடப்பெ யர்கிறது. அதே சமயம் சவ்வின் புரதங்களின் துருவத் தன்மை கொண்ட பகுதி மிக அதிகம் இருப்பதால் இவ்வியக்கத்தைச் செய்ய முடியவில்லை.
செல் சவ்வின் பணிகள்:
செல் சமிக்ஞைகளை ஏற்படுத்துதல், ஊட்டங்களை இடப்பெயரச்செய்தல், நீரைக்கடத்துதல், தேவையற்ற பொருட்கள் செல்லினுள் புகாமல் தடுத்தல் போன்ற பல்வேறு பணிகளைச் செல் சவ்வு செய்கிறது.
சைட்டோபிளாசம்:
செல்லின் பல்வேறு செயல்களுக்கு முக்கிய இருப்பிடமாக (பரப்பாக) சைட்டோபிளாசம் திகழ்கிறது. இது செல்லை நிரப்பும் ஜெலாட்டின் என்ற பகுதி திரவத்தினாலான கூழ்மமாகும். சைட்டோபிளாசம் 80% நீரால் ஆனது. இது தெளிவாகவும்மற்றும்நிறமற்றதாகவும்காணப்படும். சைட்டோபிளாசம் புரோட்டோ பிளாசத்தின் உட்கரு அற்ற பகுதி எனக் கூறப்படுகிறது. சைட்டோபிளாசம் மூலக்கூறுகள் நிறைந்த ஊட்டச்சத்து திரவமாகும். இதனுள் இரட்டை லிப்பிடுகளான (Lipid bilayer), சவ்வு சூழ்ந்த அனைத்துச் செல் உள்ளுறுப்புகள் பொதிந்துள்ளன. இதில் ஊட்டச்சத்துகள், உப்புகள் கரைந்த நிலையில் உள்ளன மேலும் கழிவுப் பொருட்களைக் கரைப்பதற்கு அமிலங்களும் காணப்படுகின்றன. இது செல் உள்ளுறுப்புகளுக்குப் பாதுகாப்பளிக்கிறது. செல் உட்பொருட்கள் செல்லைச் சுற்றி நகர இதில் நிகழும் சுழல் ஓட்டம் உதவுகிறது. சைட்டோபிளாசத்தில் பல உப்புகள் நிறைந்திருப்பதால் சிறந்த மின்கடத்தியாகச் செயல்படுகிறது. செல்லின் பிளாஸ்மா சவ்விற்கும் உட்கரு சவ்விற்கும் இடைப்பட்ட திரவப் பகுதியே சைட்டோபிளாசமாகும். பெரும்பாலான செல் வளர்சிதை மாற்ற வழித்தடங்களான கிளைக்காலிஸிஸ் மற்றும் செல் பகுப்பு ஆகியவை சைட்டோபிளாசத்தில் நிகழ்கிறது.
செல் உறுப்புகள்:
எண்டோமெம்பிரேன் அமைப்பு:
யூகாரியோடிக் கலத்தில் உள்ள சவ்வுகளின் அமைப்பு, பிளாஸ்மா சவ்வு, அணு சவ்வு, எண்டோபிளாஸ்மிக் ரெட்டிகுலம், கோல்கி கருவி, லைசோசோம்கள் மற்றும் வெற்றிட சவ்வுகள் (Tonoplast) ஆகியவற்றை உள்ளடக்கியது. எண்டோமெம்பிரேன்கள் பாஸ்போலிப்பிட்கள் மற்றும் உட்பொதிக்கப்பட்ட புரதங்களால் ஆனது, அவை சைட்டோபிளாஸில் ஏற்படும் உயிரணு சவ்வைப் போலவே இருக்கும். எண்டோமெம்பிரேன் அமைப்பு முதல் யூகாரியோட்டுகளின் மூதாதையரில் உள்ள செல் சவ்வின் உள்நோக்கிய வளர்ச்சியிலிருந்து உருவாகிறது.
எண்டோபிளாச வலை:
உள்சவ்வுத் தொகுப்பில் மிகப் பெரிதாகக் கருதப்படுவது எண்டோபிளாச வலை ஆகும்.
இதற்கு இப்பெயரிட்டவர் K.R. போர்டர் (1948) என்ற அறிஞர் ஆவார். எண்டோபிளாச வலை இரட்டைச் சவ்வினால் ஆனது.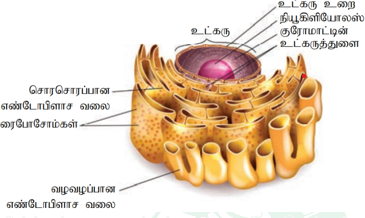
புற அமைப்பில் கீழ்க்கண்ட அமைப்புக்கூறுகளை இது பெற்றுள்ளது.
- சிஸ்டெர்னே – இது நீள, அகல மற்றும் தட்டையான பை போன்ற அமைப்புகளுடன் இணை கற்றைகளாக அமைந்த சவ் வு தொகுப்பாகும். இச்சவ்வின் குவியல் லாமெல்லா அமைப்பு போலக் காணப்படுகின்றது. சிஸ்டெர்னே சவ் வுகளில் இடைவெளி பகுதி திரவம் நிறைந்த பகுதியாக உள்ளது.
- வெசிக்கிள்கள் – முட்டை வடிவ, சவ்வு சூழ்ந்த உட்குமிழ்கள் வெசிக்கிள்கள் ஆகும்.
- டியூபியூல்கள் – இவை ஒழுங்கற்ற கிளைத்த மென்மையான சுவருடைய உள்வெளியைப் பெற்ற அமைப்புகளாகும்.
எண்டோபிளாஸ்மாச்சவ்வு மற்றும் உட்கருச் சவ்வுடன் தொடர்பு கொண்டுள்ளது. இது செல்லின் சைட்டோ பிளாசத் தினுள் ஒரு வலைப்பின்னலைப் போன்று அமைந்திருப்பதன் மூலம் செல்லிற்கு உறுதியைத் தருகின்றது. செல்லின் தேவைக்கேற்ப இதனுள் உள்ள வேதிச்சூழல், செல்லிற்குத் தேவையானபுரதங்களின் மடிப்புமற்றும் அவற்றின் பணிக்கேற்ப மாற்றங்கள் நிகழ்த்த உதவுகிறது. தவறான மடிப்பைக் கொண்ட புரதங்களை வெளியேற்றி, சிதைக்க எண்டோபிளாச வலை உதவுகிறது.
இதன் வெளிப்பரப்பில் ரைபோசோம்கள் ஒட்டிக் காணப்பட்டால் அதற்குச் சொரசொரப்பான எண்டோபிளாச வலை (RER) என்றும், ரைபோசோம் அற்று காணப்பட்டால் அதற்கு வழவழப்பான எண்டோபிளாச வலை (SER) என்றும் அழைக்கப்படுகிறது. வழவழப்பான எண்டோபிளாச வலை லிப்பிடு உருவாக்க உதவும் இடமாகவும் சொரசொரப்பான எண்டோபிளாச வலை புரதச் சேர்க்கை நிகழும் இடமாகவும் திகழ்கின்றன. தீமை விளைவிக்கும் சில வேதி சேர்மங்களையும், லிப்பிடில் கரையும் மருந்துப் பொருட்களையும், நச்சுநீக்க உதவும் நொதிகளையும் வழவழப்பான எண்டோபிளாச வலை பெற்றிருப்பது குறிப்பிடத்தக்கது.
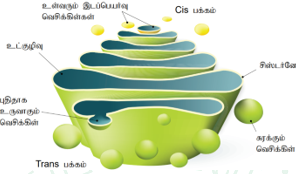
கோல்கை உடலம் (Dictyosomes):
காமிலோ கால்ஜி (1898) என்பவர் உட்கருவிற்கு அருகமைந்த வலை பின்னல் வடிவிலுள்ள இழைகளைக் கண்டறிந்தார். இந்த உள்வலை அமைப்பு பின்னர் அவரது பெயராலேயே கோல்கை உடலங்கள் என்று அழைக்கப்பட்டது. சிறிய வெசிக்கிள்களாகத் தாவரங்களில் காணப்படும் இவை டிக்டியோசோம்கள் என அழைக்கப்படுகின்றன. கோல்கை உடலமானது தட்டையான சவ்வு சூழ்ந்த பை கள் போன்ற அமைப்பாகும். இவை சிஸ்டர்னே, டியூபியூல்கள், வெசிக்கிள்கள் மற்றும் கோல்கை வாக்குவோல்களை கொண்டுள்ளன. தாவரங்களில் சிஸ்டெர்னே 10-20 எண்களைக் கொண்ட குவியல்களாகக் காணப்படுகிறது. இந்தக் குவியல்கள் ஒவ்வொன் றும் மெல்லிய அடுக்காகச் சைட்டோபிளாசத் தில் காணப்படுகிறது. சிஸ்டெர்னேவின் வெளிவிளிம்பு வலைப்பின்னலுடைய டியூபியூல்கள் மற்றும் வெசிக்கிள்களை கொண்டுள்ளது.
டியூபியூல்கள் சிஸ்டெர்னேயை ஒன்றுடன் ஒன்று இணைக்கின்றன. இவற்றின் விட்டம் 30-50nm விட்டம் ஆகும். வெசிக்கிள்கள் பெரிய உருண்டையான அல்லது குழிவுபெற்றபை போன்று காணப்படுகிறது. டியூபியூல்களின் விளிம்பிலிருந்து சிறு பைகள் போன்று வெசிக்கிள்கள் தோன்றுகின்றன. இவை வழவழப்பாகச் சுரக்கும் தன்மையுடன் காணப்படுகிறது. கோல்கை வாக்குவோல்களில் சில பெரிய உருண்டையான துகள்கள் நிறைந்த அல்லது உருவில்லாத பொருட்களைக் கொண்டு காணப்படுகிறது. இவற்றில் சில லைசோசோம் போன்று பல பணிகளை மேற்கொள்கிறது. செயலாக்கப் புரதங்களை உருவாக்க உதவும் தொடர் நிலைகளைத் தனித்தனியே நிகழ்த்தக் கோல்கை உடலங்கள் உதவுகின்றது.
சொரசொரப்பான எண்டோபிளாச வலையின் விளிம்பிலிருந்து சிறு பைகள் போன்றஅமைப்புகள் வெளியே சிறிய வெசிக்கிள்களாக மாறுகின்றன. இவ்வகையான வெசிக்கிள்கள் இணைந்து கோல்கை உடலம் தோற்றுவிக்கப்படுகின்றன. லிப்பிடுகளில் கிளைகோஸைல் ஏற்றமடையச் செய்யவும், புரதம் மொழிபெயர்பிற்குப் பின் புரத மூலக்கூறுகளில் மாற்றங்கள் நிகழவும் கோல்கை உடலங்கள் உதவுகின்றன.
பணிகள்:
- கிளைக்கோபுரதங்கள் மற்றும் கிளைக்கோலிப்பிடுகளைத் தயாரித்தல்
- லிப்பிடுகளைக் கடத்துதல் மற்றும் சேமித்தல்
- லைசோசோம்களை உருவாக்குதல்.
- செரிமான நொதிகளை உருவாகுதல்.
- செல் தட்டு மற்றும் செல் சுவரை உருவாக்குதல்
- தாவரச் செல் சுவர் ஆக்கத்திற்கும், பூச்சிகளில் கியூட்டிகள் ஆக்கத்திற்கும் உதவும் கார்போஹைட்ரேட்டுகளை சுரக்கிறது.
- ஜிமோஜென் துகள்களை (நொதிகளின் முன்னோடிகள்) உருவாக்குதல்.
மைட்டோகாண்டிரியா:
மைட்டோகாண்டிரியத்தை முதன்முதலாகக் A.கோலிக்கர் கண்டறிந்தார். இவைகளைப் பையோபிளாஸ்டுகள் என்று ஆல்ட்மேன் பெயரிட்டார். பின்னர் பெண்டா இவைகளை மைட்டோகாண்டிரியங்கள் என்று பெயரிட்டார். இவை முட்டை, உருண்டை, கோள் வடிவிலோ அல்லது செல்லின் செயல் நிலைக்கேற்ப வடிவத்தை மாற்றிக்கொள்ளும் அமைப்பாகக் காணப்படுகிறது. இது வெளி சவ்வு மற்றும் உள்சவ்வு ஆகிய இரட்டைச் சவ்வினால் ஆனது.
வெளி சவ்வானது சிறு மூலக்கூறுகளைத் தன்னுள் செலுத்தும் மென்மையான சவ்வாக உள்ளது. இதில் போரின்கள் என்ற புரதங்கள் காணப்படுகின்றன. இவை கால்வாய் போன்று அமைந்து 1000 டால்டனுக்கும் சிறிதாக உள்ள மூலக்கூறுகளைத் தம்முள் செலுத்தும் தன்மையுடையவை. மைட்டோகாண்டிரியத்தின் உள்சவ்வு, மைட்டோகாண்டிரியத்தை இரண்டு அறைகளாகப் பிரிக்கின்றது. வெளி அறையானது இரண்டு சவ்விற்கு இடையில் காணப்படுகிறது. இதற்கு மைட்டோகாண்டிரிய புறவெளி என்றும் உள் அறை மாட்ரிக்ஸ் என்ற பொருளால் நிரப்பப்பட்டுள்ளது.
உள்சவ்வு உட்புறமாக மடிப்புகளை உருவாக்குகின்றன. இந்த மடிப்பு நீட்சிகளுக்குக் கிரிஸ்டே என்று பெயர். எலக்ட்ரான் கடத்து அமைப்பின் பல நொதிகள் கிரிஸ்டேவில் காணப்படுகிறது. இதன் உள்அறை புரதப் பொருளாலானது. இதற்கு மைட்டோ காண்டிரியல் மாட்ரிக்ஸ் என்று பெயர். உள்உறையின் பரப்பில் காம்பு போன்ற துகள்கள் காணப்படுகின்றன. இவை தொடக்க நிலை துகள்கள் அல்லது பெர்னான்டியா மோரன் துகள்கள், F1 துகள்கள் அல்லது ஆக்ஸிசோம்கள் என்று அழைக்கப்படுகின்றன.
ஒவ்வொரு F1 துகளும் வட்டமான தலை, காம்பு மற்றும் அடிப்பகுதி என மூன்றுப் பகுதிகளைப் பெற்றுள்ளது. இவற்றுள் தலைப்பகுதியில் ஆக்சிகரணப் பாஸ்பரிகரணத்திற்குத் தேவையான ATP சின்தேஸ் என்ற நொதி காணப்படுகிறது. பல அயனிகள், சிறுமூலக்கூறுகள் ஆகியவற்றை ஊடு கடத்த இயலாத சவ்வாக உள்சவ்வு உள்ளது. ஆக்சிகரணப் பாஸ்பரிகரணத்திற்கு உதவும் புரோட்டான் வாட்டத்தைத் தக்க வைக்க இச்சவ்வு உதவுகிறது.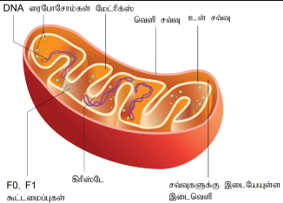
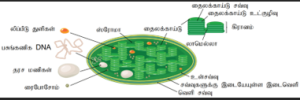
பசுங்கணிகம்:
பசுந்தாவரத்தின் அதி முக்கிய உள்ளுறுப்பாகப் பசுங்கணிகம் கருதப்படுகிறது. பசுங்கணிகம் உள்சவ்வு, வெளிசவ்வு என இரட்டைச் சவ்வினால் ஆனது. இவ்விரு சவ்வுகளுக்கிடையே உள்ள பகுதி பசுங்கணிக சுற்றுவெளி என அழைக்கப்படுகிறது. உள்சவ்வினால் சூழப்பட்ட உள்வெளியில் ஜெல்லாடினஸ் மேட்ரிக்ஸ், லிப்போபுரத திரவம் காணப்படுகின்றன. இப்பகுதிக்கு ஸ்ட்ரோமா என்று பெயர். ஸ்ட்ரோமாவினுள் தட்டையான பின்னப்பட்ட நிலையில் உள்ள பகுதிக்குத் தைலக்காய்டுகள் என்ற சவ்வு வட்டில்கள் காணப்படுகின்றன. தைலகாய்டு சவ்வு தைலக்காய்டு உள்வெளியைச் சூழ்ந்துள்ளது.
பல தைலகாய்டுகளின் தொகுப்பு கிரானம் எனப்படுகிறது. இது ஒன்றன் மேல் ஒன்றாக அமைந்து நாணய அடுக்கு போல் காணப்படுகிறது. சூரிய ஒளியின் ஆற்றலை ஈர்த்துக் கிரானங்கள் அதை வேதிய ஆற்றலாக மாற்றுகின்றன. இந்த வேதிய ஆற்றலைக் கொண்டு ஸ்ட்ரோமா பகுதி கார்போஹைட்ரேட்டுகளைத் தயாரிக்கிறது. தைலகாய்டுகளில் பச்சைய நிறமி காணப்படுகிறது. பசுங்கணிகங்களில்ஆஸ்மிய ஈர்ப்பு திறன் கொண்ட சிறுதுகள்கள், 70S ரைபோசோம்கள், DNA (வட்ட வடிவம் மற்றும் ஹிஸ்டோன்கள் அற்றவை) மற்றும் RNA ஆகியவை காணப்படுகின்றன. ஒளிச்சேர்க்கையில் பங்காற்றும்சுமார் 30புரதங்கள், ஒளி அமைப்பு I மற்றும் ஒளி அமைப்பு II சைட்டோகுரோம் bf தொகுப்பு, ATP சின்தேஸ் நொதி உருவாக்க, பசுங்கணிகத்தின் ஜீனோம் குறியீடு உதவுகிறது. Rubisco நொதியின் ஒரு துணை அலகு பசுங்கணிகத்தின் DNA-வால் குறியீடு செய்யப்படுகிறது. பசுங்கணிகத்தின் ஸ்ட்ரோமாவில் காணப்படும் முக்கியமான புரதமாக Rubisco கருதப்படுகிறது. உயிரி உலகின் அதிகம் காணப்படும் புரத மூலக்கூறாக Rubisco உள்ளது. தைலகாய்டுகளில் உள்ள சிறிய, வட்ட வடிவமான ஒளிச்சேர்க்கை அலகுகளுக்குக் குவான்டசோம்கள் என்று பெயர். பசுங்கணிகங்களும் “பாதி தற்சார்புடைய செல் நுண்ணுறுப்பாக” உள்ளன. இவைகளும் மைட்டோகாண்டிரியங்களைப் போலப் பிளவுறுதல் மூலம் பெருக்கமடைகின்றன.
பணிகள்:
- ஒளிச்சேர்க்கை
- கிரானாவில் ஒளிச்செயலை நிகழ்த்துதல்
- ஒளி சார்ப்பற்ற வினைகளை ஸ்ட்ரோமாவில் நிகழ்த்துதல்
- ஒளி சுவாசத்தில் பங்காற்றுதல்
ரைபோசோம்கள்:
ரைபோசோம்களை முதலில் கண்டறிந்தவர் ஜார்ஜ் பாலேடு ஆவார். இவைகளை செல்லில் மிக அதிகச் செறிவுள்ள துகள்கள் அல்லது மணிகளாக மின்னணுநுண்ணோக்கியின் மூலம் கண்டறிந்தார். ரைபோசோம்கள் ஒவ்வொன்றும் பெரியதும், சிறியதுமான இரு துணை அலகுகளைக் கொண்டுள்ளதாக மின்னணு நுண்ணோக்கி மூலம் கண்டறியப்பட்டது. இவ்விரு துணை அலகுகளும் ஒட்டியிருப்பது Mg2+ செறிவைப் பொருத்தது. ரைபோசோம்கள் தானாக நியூக்ளியோலஸ் பகுதியிலிருந்து தான் தோன்றியதாகவும், சுயமாக இரட்டிப்படையும் அமைப்புகளாகவும் திகழ்கின்றன. ஒவ்வொரு ரைபோசோமும் பெரிய மற்றும் சிறிய அலகுகள் என இரு துணை அலகுகளைக் கொண்டது. புரதச் சேர்க்கை நிகழும் இலக்குகளா க ரைபோ சோம்கள் திகழ்கின்றன. மேலும் இவைகள் சவ் வு சூழா அமைப்புகளாக உள்ளன.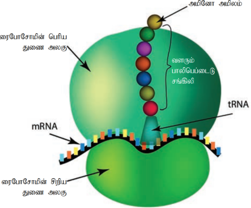
ரைபோசோம்கள் RNA மற்றும் புரதத்தால்ஆனது. இதில் RNA 60% மற்றும் புரதம் 40% அடங்கும். புரதச் சேர்க்கையின் போது பல ரைபோசோம்கள் ஒரு தூதுவ RNA (mRNA)வினால் பிணைக்கப்படுகின்றன. இதனால் தோன்றும் ஒரு கூட்டு அமைப்பிற்குப் பாலிசோம்கள் அல்லது பாலிரைபோசோம்கள் என்று பெயர்.
இந்தப் பாலிசோம்களின் பணியானது புரதச் சேர்க்கையின் போது பல நகல்களைக் கொண்டகுறிப்பிட்ட பாலிபெப்டைடுகளை உருவாக்குவதாகும். ரைபோசோமின் இரு துணை அலகுகளும் புரதச்சேர்கை நிகழாத செல்களில் தனி அலகுகளாகவும், புரதச்சேர்க்கை நிகழும் செல்களில் Mg2+ அயனியால் பிணையுற்ற அலகுகளாகவும் காணப்படுகின்றன.
ரைபோசோம்களின் வகைகள்:
70 S ரைபோசோம்கள் (துணை அலகு 30 S மற்றும் 50 S)
3 RNA மூலக்கூறுகள்
- 30 S துணை அலகு 16 SrRNA
- 50 S பெரிய துணை அலகு 23 S மற்றும் 5 S
(புரோகேரியோட்டிக் செல்களான நீலப்பச்சை பாசிகள், பாக்டீரியங்கள்; மைட்டோகாண் டிரியங்கள் மற்றும் பசுங்கணிகங்கள் கொண்ட பல பாசிகள் மற்றும் உயர்த் தாவரங்கள்)
லைசோசோம்கள் (செல்லின் தன்னைத்தானே அழித்துக் கொள்ளும் நுண்ணுறுப்பு):
லைசோசோம்களை கிரிஸ்டியன் டி டுவி (1953) கண்டறிந்தார். இவை தன்னைத்தானே அழித்துக் கொள்ளும் நுண்ணுறுப்பு என்றும் அழைக்கப்படுகிறது. இவை கோளவடிவம் கொண்டு ஒற்றைச்சவ்வால் ஆன அமைப்புகளாக யூகேரியோட்டிக் செல்களில் காணப்படுகின்றன. கோல்கை உடலத்தின் முனை சிறு பைகளாகப் பிதுக்கப்பட்டு வெளியேறும் சிறிய வாக்குவோல்கள் லைசோசோம்களாக உருவாகின்றன.
லைசோசோம்களில் காணப்படும் பல நீராற்பகுப்புற செய்யும் நொதிகள் செல்லினுள் பொருட்களைச் செரிக்க உதவுகிறது. லைசோசோம்களைச் சூழ்ந்துள்ள சவ்வு இந்த நொதிகளால் லைசோசோம்கள் அழிந்து போவதைத் தவிர்க்கிறது.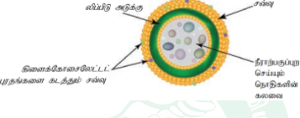
பணிகள்:
- செல்லிற்குள்ளே செரித்தல்: சைட்டோபிளாசத்தில் காணப்படும் கார்போஹைட்ரேட்டுகள், புரதங்கள் மற்றும் லிப்பிடுகளைச் செரித்தல்.
- சுய அழிவு (Autophagy): சில சாதகமற்ற சூழ்நிலையில் தன்னுடைய செல் நுண்ணுறுப்புகளான மைட்டோகாண்டியங்கள் மற்றும் எண்டோபிளாச வலை போன்றவற்றைச் செரிக்கச் செய்தல்.
- சுயச் சிதைவு (Autolysis): நோயுற்ற செல்களைச் சிதைத்துச் செல் அழிவை ஏற்படுத்துதல்.
- முதுமையடைதல் (Ageing): செல்லின் உட்புறத்தில் காணப்படும் மூலக்கூறுகளைச் சுயச் சிதைவைச் செய்யும் நொதிகளைப் பெற்றிருத்தல்.
- உள் விழுங்கும் செயல் (Phagocytosis): பெரிய செல்கள் அல்லது உட்பொருட்களைப் ஃபேகோசைட்டோசிஸ் உள்விழுங்கி செரித்துப் ஃபேக்கோசோம்-ஐ சைட்டோபிளாசத் தினுள் உருவாக்குகிறது. இந்தப் ஃபேக்கோசோமானது லைசோசோமுடன் இணைந்து செரித்தலில் பங்குகொள்கிறது.
- புறத்தள்ளல் (Exocytosis): லைசோசோம்களின் நொதிகள் செல்லிலிருந்து வெளியேற்றப்பட்டுச் செல்லின் வெளியில் உள்ள மற்ற செல்களைச் சிதைவடைய செய்தல்.
நுண் உடலங்கள்:
யூகேரியோட்டிக் செல்களில் நொதிகள் பலவற்றைப் பெற்ற சவ்வுசூழ்ந்த நுண் வெசிக்கிள்கள் நுண் உடலங்கள் எனப்படுகின்றன. இது ஒற்றைச் சவ்வினைக் கொண்ட செல் நுண்ணுறுப்பாகும். எடுத்துக்காட்டு பெராக்சி சோம்கள் மற்றும் கிளையாக்சிசோம்கள்.
பெராக்சிசோம்கள்:
பெராக்சிசோம்களை செல் நுண்ணுறுப்புகள் என்று கண்டறிந்து விளக்கியவர் கிரிஸ்டியன் டி டுவி (1967). இது சிறிய வட்ட வடிவ, ஒற்றைச் சவ்வினால் சூழப்பட்ட அமைப்பாகும். பெராக்சிசோம்கள் ஒளிசுவாசம் மற்றும் கிளைக்கோலேட் வளர் சிதை மாற்றத்தில் பங்காற்றுகிறது. தாவர இலை செல்களில் பல பெராக்சிசோம்கள் காணப்படுகின்றன. பாலூட்டிகளின் கல்லீரல், சிறுநீரகம், புரோடோசோவன்கள், ஈஸ்ட் செல்கள் ஆகியவற்றில் இவை அதிகம் காணப்படுகின்றன.
கிளையாக்ஸிசோம்கள்:
கிளையாக்ஸிசோம்களைக் கண்டறிந்தவர் ஹாரி பிவேர்ஸ் (1961). இவை தாவரச் செல்களில் மட்டும் காணப்படும் ஒற்றைச் சவ்வைக் கொண்ட, துணை செல் நுண்ணுறுப்பாகும். இது கிளையாக்சிலேட் வழித்தடத்திற்குத் தேவையான நொதிகளைக் கொண்டுள்ளது. முளைக்கும் விதைகளில் காணப்படும். கிளையாக்சிசோம்கள், கொழுப்பு அமிலங்களின் ஆக்சிகரணம் நிகழ உதவுகின்றன. எடுத்துக்காட்டு: ஆமணக்கு விதைகள்.
ஸ்ஃபீரோசோம்கள்:
இவை கோள வடிவம் கொண்டு, ஒற்றைச் சவ்வினால் சூழப்பட்ட நுண்ணுறுப்புகள் ஆகும். எடுத்துக்காட்டு எண்ணெய் வித்துகளில் உள்ள கருவூண் செல்களில் கொழுப்புப் பொருளைச் சேமித்தல்.
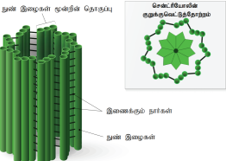
சென்ட்ரியோல்கள்:
டியூபியூலின் என்ற பொருளால் ஆன மூன்றின் தொகுப்பாக விளங்கும் ஒன்பது புறநுண் இழைகள் இவைகளாகும். சென்ட்ரியோலின் மையப்பகுதிக்கு ஹப் என்று பெயர். இதிலிருந்து ஆரப்போக்கில் விரியும் ஆரைகள் வெளிப்புற மூவிழைகளுடன் இணைந்துள்ளன (9 + 0 முறை).
குறுயிழைஅல்லதுகசையிழைகள் மற்றும்விலங்கு செல்லில் கதிர்கோல் இழைகள் தோற்றுவிக்கும் கதிர்கோல் சாதனம் ஆகியவற்றை உருவாக்கும் அடி உடலங்களாகச் சென்ட்ரியோல்கள் திகழ்கின்றன. இவை சவ்வினால் சூழப்படாத செல் நுண்ணுறுப்புகளாகும்.
வாக்குவோல்கள்:
தாவரச் செல்களில் வாக்குவோல்கள் பெரிதாகவும், டோனோபிளாஸ் ட்டு என்ற ஒற்றைச் சவ்வினால் சூழப்பட்டும் காணப்படுகிறது. அதன் செல் சாற்றில் சர்க்கரை, அமினோ அமிலங்கள், கனிம உப்புகள், வேதிய கழிவுப் பொருட்கள் மற்றும் ஆந்தோசையானின் நிறமிகள் கரைந்த நிலையில் காணப்படுகிறது. பீட்ரூட் செல்களின் வாக்குவோல்களில் ஆந்தோசையானின் நிறமி அதிகம் உள்ளது. டானின் பொ ருட்கள் செல்லில் சேகரம் அடைய இவை உதவுகின்றன. சவ்வூடு பரவல் மூலம் நீர் செல்லைச் சென்றடைய வாக்குவோல்கள் உதவுகின்றன. பிளாஸ்மா ச் சவ்வு சிதைவடைந்த செல்களை நீரில் இடும்போது அவற்றுள் சவ்வூடு பரவல் மூலம் நீர் உட்செல்வதை ஒழுங்குபடுத்த இவை உதவுகின்றன. தாவர வாக்குவோல்களின் முக்கியப் பணியானது நீரின் அழுத்தமான விறைப்பு அழுத்தத்தை நிலைநாட்டச் செய்வதாகும். இச்செயல் தாவர வடிவுருவத்தைக் கட்டமைக்க உதவுகிறது. இவ்வாறு பொருட்களைச் சேமிக்கும் மற்றும் ஒதுக்கம் அடையும் இலக்குகளாக வாக்குவோல்கள் திகழ்கின்றன.
எடுத்துக்காட்டு: செல்லில் உள்ள பெரும்பலான சுக்ரோஸ் சேர்மங்கள் தாவர வாக்குவோல்களில் சேமிப்புப் பொருளாகக் காணப்படுகிறது.
- கரும்பு மற்றும் பீட்ரூட் தாவரங்களில் சர்க்கரையைச் சேமித்தல்.
- ஆப்பிள் கனிகளில் மாலிக் அமிலத்தைச் சேமித்தல்.
- சிட்ரஸ் கனிகளின் செல்களில் அமிலங்களைச் சேமித்தல்.
- ஆன்டிரைனம் மலர்களின் அல்லி இதழ்களில் ப்ளேவோனாய்டு நிறமியான சையனிடின் 3 ருட்டினோசைட்டுகளை சேமித்தல்.
உட்கரு (Nucleus):
செல்லினுள் காணப்படும் முக்கியமான நுண்ணுறுப்பு உட்கரு ஆகும். இது செல்லின் அனைத்துச் செயல்களையும் கட்டுப் படுத்துகின்றது. உட்கரு பாரம்பரியப் பண்புகள் வெளிப்படக் காரணமாகவுள்ளது. இவை மற்ற செல் நுண்ணுறுப்புகளைக் காட் டிலும் அளவில் பெரியவை. இது கோளம், கனசதுரம், பலகோணம் அல்லது தட்டு வடிவினைப் பெற்றுள்ளன.
இது உள் மற்றும் வெளிஎனஇரட்டைச் சவ்வினால் ஆன உட்கரு உறையைக் கொண்டுள்ளது. உள் சவ்வானது ரைபோ சோம்கள் அற்று மென்மையாகக் காணப் படுகிறது. வெளி சவ்வானது ரைபோசோம்கள் பெற்றுச் சொரசொரப்பாக, ஒழுங்கற்ற இடைவெளிகளில் எண்டோபிளாச வலையுடன் தொடர்பு கொண்டு காணப்படுகிறது. இதன் சவ்வில் பல துளைகள் உள்ளன. இவற்றிற்கு உட்கருத்துளைகள் என்று பெயர். இந்தத் துளை மூலம் mRNA, ரைபோசோமல் அலகுகள், புரதங்கள் மற்றும் பிற பெரிய மூலக்கூறுகள் பரிமாற்றம் அடைகின்றன. ஒவ்வொரு துளையும் அனுலஸ் என்ற ஒரு வட்ட அமைப்பினால் சூழப்பட்டுள்ளது. துளையும், அனுலசும் சேர்ந்து துளை கூட்டமைப்பு ஆகும். இரண்டு சவ்விற்கும் இடையே உள்ள இடைவெளிக்கு உட்கரு புறவெளி என்று பெயர்.
உட்கரு உள்வெளியில் உள்ள ஜெலாட்டினஸ் மாட்ரிக்ஸ் உட்கருபிளாசம் என அழைக்கப்படுகின்றனது. மாட் ரிக்ஸில் செறிவற்றுப் பரவலாகக் காணப்படும் குரோமாட்டின் வலைபின்னல்களும், நியூக்ளியோலஸ்சும் இடம் பெற்றுள்ளன. இடைக்கா லச் செல் பிரிதல் நிலையில் வலைப்பின்னல்களைக் கொண்டகுரோமாட் டினானது, சுருள்களற்ற இழைகளாகக் காணப்படுகிறது. யூகேரியோட்டிக் செல்களின் குரோமாட் டின்களில் சிறிதளவு RNA மற்றும் DNA-யுடன் இணைந்த ஹிஸ்ட்டோன் புரதங்களும் உள்ளன.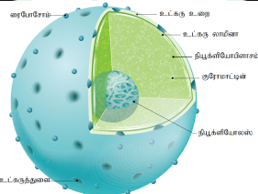
செல் பகுப்பின் போது குரோமாடின்களின் சுருக்கமடைந்த அமைப்பிற்குக் குரோமோசோம்கள் என்று பெயர். யூகோரியோட்டிக் குரோமோசோமின் பகுதியானது m-RNAவாக படியெடுக்கையில் அதில் உள்ள செயல்படும் ஜீன்கள் இடைக்கால நிலையில் இருப்பதில்லை. இதற்கு யூகுரோமாட்டின் என்று பெயர். இடைக்கால நிலையில் யூகேரியோட்டிக் குரோமோசோமின் பகுதி m-RNA வில் படியெடுக்கப்படாமல், சுருங்கி அமைந்து அதிக சாயம் ஏற்கும் பகுதி ஹெட்டிரோகுரோமாட்டின் என அழைக்கப்படுகிறது. உட்கருவினுள் ஒன்று அல்லது பல எண்ணிக்கைகளில் காணப்படும் சிறிய செறிவுற்ற கோள வடிவச் சவ்வு சூழ்ந்திராத அமைப்புகள் நியூக்ளியோலஸ் எனப்படுகின்றன. rRNA மற்றும் tRNA உருவாக்கத்திற்குத் தேவையான ஜீன்களை நியூக்கிலியோலஸ் பெற்றுள்ளது.
உட்கருவின் பணிகள்:
- செல்லின் செயல்கள் அனைத்தையும் கட்டுப்படுத்துதல்.
- மரபு அல்லது பாரம்பரியச் செய்திகளைச் சேமித்து வைத்தல்.
- புரதங்கள் மற்றும் நொதிகள் உருவாவதற்குத் தேவையான மரபுச் செய்தியை DNA-யில் பெற்றிருத்தல்.
- DNA இரட்டிப்பாதல் மற்றும் படியெடுத்தல் நிகழ்வுகளை நடத்துதல்.
- நியூக்ளியோலஸ்சில் ரைபோசோம்கள் தோன்றுதல்.
குரோமோசோம்கள்:
ஸ்டிராஸ்பர்கர் 1875 ஆம் ஆண்டு முதன்முதலில் யூகேரியோட்டு செல்களில் குரோமோசோம் இருப்பதைக் கண்டறிந்தார். வால்டேயர் (1888) குரோமோசோம் என்ற சொல்லை முதன்முறையாக அறிமுகப்படுத்தினார். குரோமோசோம்கள் ஜீன்களைக் கொண்டுள்ளன என்பதை முதன் முதலாகப் பிரிட்ஜஸ் (1916) என்பவர் உறுதி செய்தார். இவை DNA மற்றும் DNA சார்ந்த புரதங்களால் ஆனவை.
குரோமோசோமின் அமைப்பு:
குரோமோசோம் நூல் போன்ற நுண் இழைகளால் ஆனது. இதற்குக் குரோமாட்டின் என்று பெயர். இந்தக்குரோமாட்டின் DNA, புரதம், RNA ஆகியவற்றால் ஆனது. ஒவ்வொரு குரோமோசோமிலும் இரு ஒத்த அமைப்புகள் காணப்படுகின்றன. அவைகளுக்குக் குரோமாட்டிட்கள் என்று பெயர். இவை இரண்டும் ஒத்த அமைப்பைப் பெற்றிருப்பதால் சகோதரி குரோமாட்டிட்கள் எனக் கருதப்படுகின்றன. செல் பகுப்பின் போது குரோமாட் டிட்கள் சரியான அளவு மற்றும் வடிவத்தைக் கொண்ட ஒழுங்கமைவுடைய குரோமோசோம்களா கின்றன. இயல்பான குரோமோசோம் ஒன்றில் காணப்படும் குறுகிய பகுதிக்கு இறுக்கங்கள் என்று பெயர். இறுக்கங்கள் இரண்டு வகைப்படும் அவை முதலாம் நிலை இறுக்கம் மற்றும் இரண்டாம் நிலை இறுக்கம் எனப்படும். முதல் நிலை இறுக்கத் தில் சென்ட்ரோமியர் மற்றும் கைனிட்டோகோர் காணப்படுகிறது. இரண்டு குரோமாட்டிட்டுகளும் சென்ட்ரோமியர் பகுதியில் இணைவு பெற்றுள்ளன. சென்ட்ரோமியரின் எண்ணிக்கை சிற்றினத்திற்கு ஏற்றவாறு மாறுபடுகின்றது. மானோசென்ட்ரிக் குரோமோசோமில் ஒரு சென்ட்ரோமியரும், பாலிசென்ட்ரிக் குரோமோசோமில் பல சென்ட்ரோமியர்களும் காணப்படுகின்றன. சென்ட்ரோமியர்களில் காணப்படும் புரதஇழைகளின் கூட்டமைப்பு கைனிட்டோகோர் என்று அழைக்கப்படுகிறது. மைட்டாசிஸ் செல் பகுப்பின் போது குரோமோசோமில் காணப்படும் கைனிட்டோகோர் பகுதியில் கதிர்கோல் இழைகள் இணைக்கப்படுகின்றன.
முதலாம் நிலை இறுக்கம் மட்டுமல்லாமல் சில குரோமோசோம்களில் இரண்டாம் நிலை இறுக்கமும் காணப்படுகின்றன. இரண்டாம் நிலை இறுக்கத்தில் இருந்து உருவாகும் நியூக்ளியோலஸ்களை நியூக்ளியோலார் அமைப்பான்கள் எனப்படுகின்றன. ஒரு சில இரண்டாம் நிலை இறுக்கத்தில் ரைபோசோமல் RNA உருவாவதற்கான ஜீன்கள் காணப்படுகின்றன. இவைகள் நியூக்ளியோலஸ் உருவாவதைத் தூண்டுகின்றன. அவைகளுக்கு நியூக்ளியோலஸ் அமைக்கும் பகுதிகள் எனப்படுகின்றன.
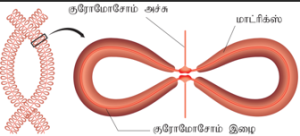
பிரதானக் குரோமோசோமின் முனைகளிலிருந்து தோன்றும் சிறு குரோமோசோம் பகுதிக்குச் சாட்டிலைட் அல்லது SAT என்று பெயர். இந்தச் சாட்டிலைட் பகுதி ஒரு நீண்ட இரண்டாம் நிலை இறுக்கத்தின் மூலம் பிரதானக் குரோமோசோமில் இருந்து பிரிகிறது. சாட்டிலைட்டுகள் காணப்படும் குரோமோசோமிற்கு SAT குரோமோசோம் என்று பெயர். சிலகுரோமோசோம்களின் உருஅமைப்பைத் தீர்மானிக்கும் பகுதியாகச் சாட்டிலைட் திகழ்கிறது.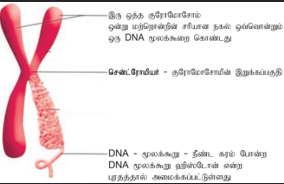
குரோமோசோம்களின் நுனி பகுதி டீலோமியர் என அழைக்கப்படுகிறது. இது குரோமோ சோமிற்கு நிலைத்தன்மை அளிக்க உதவுகிறது. இப்பகுதியில் உள்ள DNA குறிபிட்ட வரிசையில் அமைந்த நியூக்ளியோடைடுகளால் ஆனது. அனைத்து யூகேரியோட்டு குரோமோ சோம்களிலும் இப்பகுதியில் 5’TTAGGG3’ என்ற வரிசையில் பல படிகளில் அமைந்த சிறிய DNA நியூக்ளியோடைட் வரிசைகள் காணப்படுகின்றன. எடுத்துக்காட்டு: நியூரோஸ்போரா கிராசா மற்றும் மனிதர்கள். இந்த டிலோமியர் பகுதிகளை தக்கவைப்பதால், செல்லினுடைய வாழ் நா ட்காலம், இனப்பெருக்கத் தகுதி ஆகியவைத் தீர்மானிக்கப்படுகிறது. எனவே டிலோமியர் மற்றும் டிலோமிரேஸ் பற்றிய ஆய்வு வயதாதல், புற்று நோய் பற்றிய புதிய புரிதலுக்கு வழிவகுக்கும். ஒரு குரோமோசோமின் முனை மற்றொன்றுடன் பிணையுறுதலைத் தடுப்பதற்கு இந்த டிலோமியர்கள் உதவுகின்றன.
குரோமோசோம்களின் வகைகள்:
சென்ட்ரோமியரின் நிலையின் அடிப்படையில், குரோமோசோம்கள் டெலோசென்ட்ரிக் (டெர்மினல் சென்ட்ரோமியர்), அக்ரோசென்ட்ரிக் (டெர்மினல் சென்ட்ரோமியர் கேப் டெலோமியர்), சப் மெட்டாசென்ட்ரிக் (சென்ட்ரோமியர் சப்டெர்மினல்) மற்றும் மெட்டாசென்ட்ரிக் (சென்ட்ரோமியர் மீடியன்) என்று அழைக்கப்படுகின்றன. யூகாரியோடிக் குரோமோசோம் தடி வடிவமாக இருக்கலாம் (டெலோசென்ட்ரிக் மற்றும் அக்ரோசென்ட்ரிக்), எல்-வடிவ (துணை-மெட்டாசென்ட்ரிக்) மற்றும் v-வடிவ (மெட்டாசென்ட்ரிக்).
குரோமோசோம்களின் பணிகளைக் கொண்டு அவை ஆட்டோசோம்கள் மற்றும் பால் குரோமோசோம்கள் என்று பிரிக்கப்பட்டுள்ளன. ஒரு உயிரின் உடலப் பண்பைக் கட்டுப்படுத்துவதால் எல்லா உடலச் செல்களில் ஆட்டோசோம்கள் காணப்படுகின்றன. மனிதர்களில் இரட்டை மைய எண்ணிக்கை கொண்ட செல்களில் 44 குரோமோசோம்கள் ஆட்டோசோம்களும் இரண்டு பால் குரோமோசோம்களும் உள்ளன. பால் குரோமோசோம்கள் பால் நிர்ணயத்தில் பங்கு கொள்கின்றன.
சிறப்பு வகை குரோமோசோம்கள்:
சில குறிப்பிட்ட திசுக்களில் மட்டுமே இந்தச் சிறப்பு வகை குரோமோசோம்கள் காணப்படுகின்றன. இந்தச் சிறப்பு வகை குரோமோசோம்கள் அளவில் பெரிதாக காணப் படுவதால் இவற்றை அசுரக் குரோமோசோம்கள் என்று அழைக்கின்றோம். சில தாவரங்களின் கருவின் சஸ்ப ன்சார்களில் இவை காணப்படுகிறது. விலங்குகளில் காணப்படும் பாலிடீன் குரோமோசோம்கள் மற்றும் விளக்கு தூரிகை குரோமோசோம்களும் அசுரக் குரோமோசோம்களே ஆகும்.
பாலிடீன் குரோமோசோம்கள்:
E.G. பால்பியானி என்பவர் டேராசோஃபைலா என்ற பழப் பூச்சியின் உமிழ்நீர் சுரப்பில் இதனைக் கண்டறிந்தார். இது பல்வேறு பூச்சிகளின் லார்வாக்கள், மிட்ஜஸ்யில் காணப்படுகின்றன.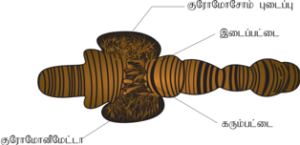
உட்கரு பகுப்பு ஏற்படாமல் இடைக்கால நிலையில் குரோமோசோம்கள் இரட்டித்தலில் ஈடுப்படுகின்றன. ஒருகுரோமோசோம் பல நகல்களை உருவாக்குவதால் தோன்றும் அமைப்பே பாலிடீன் குரோமோசோம் ஆகும். இதை ஒளி நுண்ணோக்கியிலும் காண இயலும். மரபியல் செயல்பாடு கொண்டுள்ளதாக இவை காணப்படுகின்றன. இதில் அடர்த் தியான சாயம் ஏற்கும் பட்டைகள் மற்றும் சாயம் ஏற்கா இடைப்பட்டைகள் அடுத்தடுத்துக் காணப்படுகின்றன. இவற்றுள் அடர்த் தியான பட்டையில் 95% DNA- வையும், சாயம் ஏற்கா இடைப்பட்டையில் 5% DNA-வையும் கொண்டுள்ளன. கைரோனோமஸ் லார்வாவில் உள்ள பாலிடீன் குரோமோசோம்களில் மிகப் பெரிய புடைப்புகள் காணப்படுகின்றன. இவற்றிற்குப் பால்பியானி வளையங்கள் என்று பெயர். இதற்குக் குரோமோசோம் புடைப்புகள் என்றும் அழைக்கலாம். இவை துரிதமாக RNA உற்பத்தி நிகழும் இலக்குகளாகும். குறிப்பாக இது உமிழ்நீர் சுரப்பியில் காணப்படுவதால் இவை உமிழ்நீர் சுரப்பி குரோமோசோம்கள் என்றும் அழைக்கப்படுகிறது.
உட்கரு பகுப்பு நடைபெறாமல் குரோமோசோம் DNA தொடர்ச்சியாக இரட்டிப்படைந்து தோன்றும் சகோதரக் குரோமாட்டிட்கள் பக்கவாட்டில் தொகுக்கப்பட்டு இந்தப் பாலிடீன் குரோமோசோம் உருவாகிறது. இந்த நிகழ்விற்கு எண்டோமைட்டாசிஸ் என்று பெயர். ஜீன் வெளிப்பாடு, ஜீன்கள் படியெடுத்தல் மற்றும் RNA உருவாதல் போன்றவை பாலிடீன் குரோமோசோமில் நிகழ்கின்றன. உடலச் செல்களில் தாய்வழி மற்றும் தந்தைவழித் தோன்றிய ஒத்திசைவு குரோமோசோம்கள் பக்கவாட்டில் ஒன்றொடொன்று தொடர் புடையதாகக் காணப்படுகின்றன. இதற்கு உடல இணைவு என்று பெயர்.
விளக்கு தூரிகை குரோமோசோம்கள்:
இராட்சச உட்கருவைக் கொண்ட ஒரு செல் ஆல்கா அசிடாபுலேரியா மற்றும் சலமண்டார் ஊசைட்டுகளில் முதல் மியாட்டிக் புரோஃபேஸின் டிப்லோடீன் துணை நிலையில் விளக்கு தூரிகை குரோமோசோம்கள் காணப்படுகிறது. இதைப் பிளம்மிங் (1882) முதன்முதலில் கண்டறிந்தார். அடர்த்தியான குரோமோசோம் பகுதி குரோமோசோம் அச்சை உண்டாக்கி இதில் பல நீட்சிகள் காணப்படுகிறது. இந்த நீட்சிகள் DNA-வைக் கொண்டுள்ளது. இதில் RNA உற்பத்தி நடைபெறுகிறது.
செல் சுழற்சி:
வரையறை: புதிய செல்லை உருவாக்கும் தொடர்ச்சியான நிகழ்விற்கு செல் சுழற்சி என்று பெயர். இந்த தொடர்ச்சியான நிகழ்வு பல நிலைகளைக் கொண்டுள்ளது.
பகுபடும் மனிதச் செல்லின் செல் சுழற்சி கால அளவு
நிலை | கால அளவு (மணியில்) |
G1 | 11 |
S | 8 |
G2 | 4 |
M | 1 |
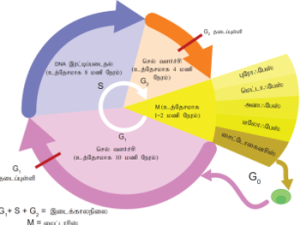
செல் சுழற்சியின் கால அளவு:
செல் சுழற்சி நிலைகளின் கால அளவு செல்களின் வகைக்கு ஏற்றவாறு வேறுபடுகிறது. யூகேரியோட்டிக் செல்லானது 24 மணி நேரத்திற்கு ஒருமுறை பகுப்படைகிறது. செல் சுழற்சியானது மைட்டாடிக் பகுப்பு நிலை மற்றும் இடைக்காலநிலை என இரண்டாகப் பிரிக்கப்படுகிறது. செல் சுழற்சியில் 95 விழுக்காடு கால அளவை இடைக்கால நிலை எடுத்துக் கொள்கிறது. மீதமுள்ள ஒரு மணி நேரம், உட்கரு பகுப்பு மற்றும் சைட்டோபிளாச பகுப்பு எடுத்துக்கொள்கின்றன.
இடைக்கால நிலை:
இடைக்கால நிலை செல் பகுப்பில் அதிகக் காலம் கொண்ட நிலை ஆகும். ஆனால் இது முற்றிலும் வேறுபட்டது என்பது குறிப்பிடத்தக்கது. பார்ப்பதற்கு உட்கரு ஓய்வு நிலையில் இருப்பது போல் தோன்றும். ஆனால் இது உண்மையல்ல. இழை போன்ற அமைப்பிலிருந்து குரோமோசோம்கள் இந்நிலையில் விரவிய அமைப்பாக உள்ளன. பெரும்பாலான நேரங்களில் இந்நிலையின் போது இவை புரத உற்பத்தியில் ஈடுபடுகின்றன.
G1 நிலை – முதல் இடைவெளி நிலை
G1 நிலையில் இருக்கும் செல்களில் DNA-வின் அளவானது 2C ஆக உள்ளது. இந்நிலையில் செல்லானது வளர்சிதை மாற்றச் செயலில் ஈடுபட்டு வளர்ச்சிக்குத் தேவையான புரதம், லிப்பிடுகள், கார்போஹைட்ரேட்டுகள் மற்றும் செல் நுண்ணுறுப்புகளான மைட்டோ காண்டிரியங்கள், எண்டோபிளாச வலை ஆகியவற்றை உருவாக்குகின்றன.
பல்வேறு தடைப் புள்ளிகள் செல் சுழற்சியைக் கட்டுப்படுத்துகின்றன. G1 படிநிலையின் முடிவில் ஏற்படும் தடைப்புள்ளி “வரையறு புள்ளி” என்று அழைக்கப்படுகிறது. ஒரு செல்லானது செல் சுழற்சியில் தொடர்ந்து செயல்படுவதை அல்லது G0 என்ற அமைதி நிலைக்குச் செல்வதை மற்றும் குறிப்பிட்ட செல்லாக மாற்றம் அடைவதை அல்லது பகுபடாமல் இறந்து விடுவதைத் தீர்மானிப்பதாக இந்தத் தடைப்புள்ளி திகழ்கிறது. G1 நிலையில் செல்கள் பகுபடாமல் தடைப டுவதற்குக் காரணம்:
- ஊட்டம் இல்லாமை.
- வளர்ச்சி ஊக்கிக்காரணிகள் இல்லாமை அல்லது செல்களின் செறிவு சார்ந்த தடை.
- வளர்சிதை மாற்றம் அடைந்து G0 நிலைக்குச் செல்கின்றது.
செல்லினுள் காணப்படும் உயிர்வேதிப் பொருட்கள் செல் பகுப்பினைச் செயல்படச் செய்கிறது. கைனேசஸ் மற்றும் சைக்ளின்கள் என்ற புரதங்கள் ஜீன்களையும் அவற்றின் புரதங்களையும் செயல்படச் செய்து செல் பகுப்பினைச் செயல்படுத்துகிறது. சைக்ளின்கள் G1 நிலையில் முக்கியத் தடைப்புள்ளியாக செயல்பட்டு ஒரு செல்லானது பகுப்படைகிறதா அல்லது பகுப்படையாமல் இருக்கின்றதா என்பதைத் தீர்மானிக்கின்றது.
G0 நிலை
சில செல்கள் G1 நிலையிலிருந்து விடுபட்டு அமைதி நிலைக்குச் செல்கின்றன. இந்நிலைக்கு G0 நிலை என்று பெயர். G0 நிலையில் செல்கள் நீண்ட காலம் செல் பெருக்க மடையாமல் இருந்து வளர்சிதை மாற்றத்தை மட்டுமே செய்கின்றன. ஆனால் பெருக்கம் அடைவதில்லை. G0 நிலையில் உள்ள செல்கள் RNA மற்றும் புரதச்சேர்க்கை செயல்களைக் குறைந்த அளவில் செய்வதுடன் வளர்ச்சியற்ற நிலையில் உள்ளன.
G0 நிலை நிலையற்றது. முதிர்ந்த நியூரான், எலும்புத்தசை ஆகியவற்றின் செல்கள் G0 நிலையில் நிலைத்துவிடுகின்றன. உகந்த செல் சாரா சமிக்ஞை மற்றும் வளர்ச்சிக் காரணிகள் கிடைத்தால் மட்டும் G0 நிலையை விட்டுப் பெருக்கமடையும் நிலைக்குப் பெரும்பாலான விலங்கினச் செல்கள் செல்ல இயலும். இல்லையெனில் G0 நிலையிலேயே நின்று விடும். G0 செல்களை வளர்வடக்க நிலையில் (Dormant) உள்ள செல்களாகக் கருதப்படுவதில்லை.
S நிலை – உருவாக்க நிலை – இடைப்பட்ட அளவுடைய DNA வை கொண்ட செல்கள்
DNA உற்பத்தியில் இருப்பதால், 2C-க்கும் 4C-க்கும் இடைப்பட்ட நிலையில் உள்ளதாக இது கருதப்படுகிறது. DNA இரட்டிப்பால் செல்லின் வளர்ச்சி தொடர்ந்து நிகழ்வதுடன் ஹிஸ்டோன் என்ற புரத மூலக்கூறுகள் உருவாக்கப்பட்டு, DNA-வுடன் இணைக்கப்படுகின்றன. சைட்டோபிளாசத்தில் சென்ட்ரியோல்கள் இரட்டிப்படைகின்றன. இறுதியில் DNA அளவானது 2C-யிலிருந்து 4C-ஆக பெருக்கமடைகிறது.
G2 நிலை – இரண்டாவது இடைவெளி நிலை – G2 மற்றும் மைட்டாசிஸ் செல்களில் 4C அளவு DNA காணப்படுதல்
புரதச் சேர்க்கை மற்றும் செல் நுண்ணுறுப்புகள் உருவாதல், மைட்டோகாண்டிரியம், பசுங்கணிகம் பகுப்படைதல் கதிர்கோல் இழைகள் உருவாதல் ஆகியவை இந்நிலையின் சிறப்புப்பண்புகளாகும். இதைத் தொடர்ந்து உட்கரு பகுப்பு, சைட்டோபிளாச பகுப்பு நடைபெறுகிறது. DNA அளவு 4C-ஆகவே உள்ளது. டியூபியூலின் புரத ஆக்கத்தின் மூலம் நுண் குழல் இழைகள் தோன்றுகின்றன. நுண்குழல் இழைகள் ஒன்று சேர்ந்து கதிர்கோல் இழைகளை உருவாக்கி உட்கரு பகுப்பைச் செயல்படுத்துகின்றன.
முதிர்ச்சியை ஊக்கப்படுத்தும் காரணிகள் (Maturation Promoting Factors – MPF) என்ற ஒரு வகை புரதம் G2 காலநிலையில் மட்டுமே உருவாக்கப்படுகின்றன. இக்காரணிகள் இடைக்காலநிலை குரோமோசோம் இழைகள் செறிவுற்று, மைட்டாடிக் குரோமோசோம்களாக உருவெடுக்க உதவுகின்றன. செல் சுழற்சியில் G1, S மற்றும் G2 நிலைகளில் DNA சிதைதல் தடைப்புள்ளி செயல்படுவது குறிப்பிடத்தக்கது.
செல் பகுப்பு:
ஏமைட்டாசிஸ் (நேர்முகப் பகுப்பு):
ஏமைட்டாசிஸ், நேர்முகப் பகுப்பு அல்லது தெளிவிலாச் செல் பகுப்பு என்றும் அழைக்கப்படுகின்றன. இப்பகுப்பில் கதிர்கோல் இழைகள் தோன்றுவதில்லை. குரோமாட்டின் பொருள் செறிவுற்று குரோமோசோம்கள் உருப்பெறுவதில்லை. இதிலும் காரியோகைனசிஸ், சைட்டோகைனசிஸ் என இரு நிலைகள் உள்ளன.
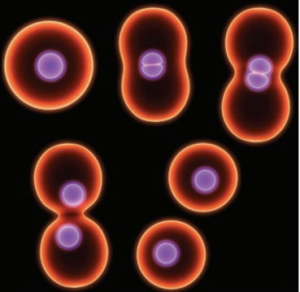
காரியோகைனசிஸ்:
- உட்கரு பகுப்படைதல்.
- உட்கருவின் இடைப்பகுதியில் இறுக்கம் ஏற்பட்டு உடுக்கை வடிவம் அடைதல்.
- இறுக்கம் ஆழமாகி உட்கரு இரண்டாகப் பிரிதல்.
சைட்டோகைனசிஸ்:
- சைட்டோபிளாசம் பகுப்படைதல்.
- உட்கரு இறுக்கத்தைத் தொடர்ந்து பிளாஸ்மாச் சவ்விலும் இறுக்கம் உருவாகுதல்.
- சவ்வில் நிகழும் இந்த இறுக்கமும் மையம் நோக்கி விரிவடைந்து (Centripetal) இறுதியில் சைட்டோபிளாசம் இரு பகுதிகளாக பிரிந்து இரு செல்கள் உருவாகுதல்.
எடுத்துக்காட்டு:
பாலூட்டிகளின் குறுத்தெலும்பு செல்களின் பகுப்பு, பரமேசியத்தின் பெரிய உட்கரு பகுப்பு, உயர்நிலை தாவரங்களில் காணப்படும் முதுமையடைந்து சிதைந்து கொண்டிருக்கும் செல்களில் நிகழும் பகுப்பு.
நேர்முகப் பகுப்பின் குறைகள்:
- குரோமோசோம்கள் சமமற்ற அளவில் சேய் செல்களைச் சென்றடைதல்.
- வளர்சிதை மாற்றம் மற்றும் இனப்பெருக்கம் ஆகியவற்றில் பிறழ்சிகள் ஏற்படுதல்.
மைட்டாசிஸ்:
செல் பகுப்பின் முக்கிய நிகழ்வுகளில் ஒன்றாக உட்கரு நிகழ்வுகள் உள்ளன. இவற்றில் மைட்டாசிஸ் உட்கரு பகுப்பு, தண்டு நுனி, வேர் நுனி, தாவரத்தின் பிற வளர் உறுப்புகளின் ஆக்குத் திசுக்களில் நடைபெறுகிறது. தாய் செல்லின் குரோமோசோம் எண்ணிக்கையை ஒத்திருப்பதால் இதற்குச் சமநிலை பகுப்பு என்று பெயர்.
மூடிய, திறந்தமைட்டாசிஸ்:
மூடிய மைட்டாசிஸ்: உட்கரு உறை சிதையாமல் குரோமோசோம்கள் எதிரெதிர் துருவங்களை நோக்கிச் செல்கின்றன. எடுத்துக்காட்டு: பல ஒற்றைச் செல் யூகேரியோட்டுகளான ஈஸ்ட் மற்றும் சளிப் பூஞ்சைகள்.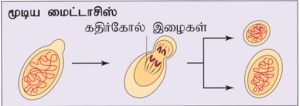
திறந்த மைட்டாசிஸ்: முதலில் உட்கரு உறை சிதைந்து. குரோமோசோம்கள் இரண்டு தொகுதிகளாகப் பிரிகிறது. பின் ஒவ்வொரு தொகுதியையும் சூழ்ந்து உட்கரு உறை மீண்டும் உருவாக்கப்படுகிறது. எடுத்துக்காட்டு: பெரும்பாலான தாவரங்கள் மற்றும் விலங்குகள்.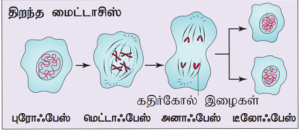
மைட்டாசிஸ் நிலைகள்:
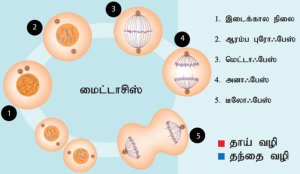
- புரோஃபேஸ்:
மைட்டாசிஸ் பகுப்பில் அதிகக்கால அளவை எடுத்துக் கொள்ளும் நிலை இதுவாகும்.
நீளமான, மெல்லிய நூல்களைப் போன்ற குரோமோசோம் அமைப்புகள் இந்நிலையில் உருவாகின்றன. செறிவுற்ற இழைகளாக உள்ள இவை மைட்டாடிக் குரோமோசோம்கள் எனப்படுகின்றன. தாவரச் செல்லில் இந்நிலையின் போதே கதிர்கோல் இழைகள் தோன்றுகின்றன. நியூக்கிளியோலஸ், உட்கருஉறை சிதைவதுடன், மறையத் தொடங்குகிறது. இந்நிலையில் கோல்கை உறுப்புகள், எண்டோபிளாச வலை ஆகியவை காணப்படுவதில்லை.
விலங்கு செல்லின் சென்ட்ரியோல்களிலிருந்து நுண் இழைகள் தோன்றிச் செல்லினுள் எதிரெதிர் துருவங்கள் நோக்கி இடப்பெயர்ச்சி அடைகின்றன. இந்த நுண் இழைகளுக்கு நட்சத்திர இழைகள் என்று பெயர். தாவரச் செல்களில் நட்சத்திர இழைகள் தோன்றுவதில்லை.
- மெட்டாஃபேஸ்
ஒரு குரோமோசோமின் சகோதரி குரோமாட்டிட்களை இணைக்கும் சென்ட்ரோமியரின் கைனட்டோகோர் பகுதியில் கதிர்கோல் இழைகள் வந்து இணைகின்றன. கதிர்கோல் இழைகள் டியூபியூலின் புரதத்தால் ஆனவை. செல்லின் மையத்தளத்தில் குரோமோசோம்கள் நெருக்கமாக அமைவதால் உண்டாகும் அமைப்பு மெட்டாஃபேஸ் தட்டு எனப்படுகிறது. இந்நிலையில் குரோமோசோமின் புறஅமைப்பு நன்கு புலப்படுகிறது. சென்ட்ரோமியரில் காணப்படும் கைனிட்டோகோர் ஆனது DNA புரதக் கூட்டுப் பொருட்களால் ஆனது. இது ஒரு மூன்று மென்தகடு வட்டத் தட்டாகக் காணப்படுகிறது.
- அனாஃபேஸ்
ஒவ்வொரு குரோமோசோமும் பிளவுற்றுப் பிரியும் இரண்டு சேய் குரோமாட்டிட்கள் செல்லின் எதிரெதிர் துருவங்களை நோக்கி இடப்பெயர்ச்சி அடைகின்றன. ஒவ்வொரு சென்ட்ரோமியரும் கதிர்கோல் இழைகள் சுருங்குவதால் பிளவுற்று, சேய் குரோமாட் டிட்கள் விடுவிக்கப்படுவதுடன் அவை துருவம் நோக்கி இடப்பெயர்ச்சி அடைகின்றன. ஒவ்வொரு பிரிவுற்ற பகுதியும் இரண்டு குரோமாட் டிட்களை பெறுகிறது (சகோதரி குரோமாட் டிட்கள் பிரிதல் அடைதல்). சகோதரி குரோமாட் டிட்களின் பிரிவு மரபு தொகையத்தின் சமப்பிரிவடையும் நிகழ்வாக இதன் மூலம் முற்று பெறுகிறது.
கதிர் இழை தொகுப்பு தடை இலக்கும், அனாஃபேஸின் பிரிநிலை அடைதலும் APC/C என்ற திரள் புரதம் ஒட்டிணைவு புரதங்களைச் சிதைக்கத் தூண்டிக் கதிர்கோல் இழைகளைச் சுருங்கச் செய்கிறது. எனவே தான் குரோமாட்டின்களானது செல் பகுப்பில் இரு துருவங்களை நோக்கி நகர முடிகிறது. எனவே செல் மெட்டாஃபேஸ் நிலையிலிருந்து அனாஃபேஸ் நிலைக்கு முன்னேறுவதற்கு உதவுகிறது.
- டீலோஃபேஸ்
சேய் குரோமோசோம்கள் இரு தொகுதிகளாகப் பிரிவுற்று எதிரெதிர் துருவங்களை அடைகின்றன. அத்துடன் கதிர்கோல் இழைகள் மறைகின்றன. இத்துடன் மரபுப்பொருளின் பகுப்பான காரியோகைனசிஸ் (உட்கரு பகுப்பு) முடிவுறுகிறது. இதனைத் தொடர்ந்து சைட்டோகைனசிஸ் (சைட்டோபிளாச பகுப்பு) ஏற்படுகிறது. நியூக்ளியோலஸ் மற்றும் உட்கரு சவ்வு மீண்டும் உருவாகிறது. ஒவ்வொரு சகோதரிக் குரோமாட்டிடுகளின் தொகுப்பைச் சூழ்ந்து உட்கரு சவ்வு தோன்றியவுடன் அவை குரோமோசோம்கள் என அழைக்கப்படுகின்றன. ஒவ்வொரு குரோமாட்டிட்டும் அதற்கென ஒரு சென்ட்ரோமியரை பெற்றுள்ளது. பின்னர் இந்தக் குரோமோசோம்கள் மெல்லிய நூலிழைகள் போலாகின்றன. தாவரச் செல் பகுப்பின் போது இரண்டு சேய் செல்களுக்கும் இடையே பிராக்மோபிளாஸ்டுகள் உருவாகின்றன. இவற்றின் இணைவால் செல் தட்டு தோன்றி இரு சேய் செல்கள் உருவாக்கப்படுகின்றன. பிரியும் இந்த இரு புதிய சேய் செல்களிலும் பெரு மூலக்கூறுகள் மற்றும் செல் நுண்ணுறுப்புகள் பகிர்ந்தளிக்கப்பட்டு முழுமை பெற்ற சேய் செல்கள் உருவாகின்றன.
சைட்டோகைனசிஸ்:
விலங்கு செல்களில் சைட்டோகைனசிஸ்:
விலங்கு செல்களில் இது பிளாஸ்மாச் சவ்வு சுருங்குவதால் நடைபெறுகிறது. பிளாஸ்மா சவ்வினால் ஏற்படும் சுருங்கு வளையம் ஆக்டின் மற்றும் மையோசின் சேர்ந்த நுண் இழைகளால் ஆனது. இந்த இழைகள் உள்நோக்கிச் சுருங்க உதவும் விசை ஒன்று தோன்றி இறுதியில் சைட்டோபிளாசம் இரு சம அளவில் சவ்வினால் பிரிக்கப்படுகிறது.
தாவரச் செல்லில் சைட்டோகைனசிஸ்:
டீலோஃபேஸ் நிலையில் சைட்டோபிளாசம் பிரியத் துவங்குகிறது. தாவரங்களில் செல் தட்டு மையப்பகுதியில் தொடங்கி வெளி நோக்கி நகர்ந்து (Centrifugal) பக்கவாட்டில் செல்சுவரை அடைகிறது. பிராக்மோபிளா ஸ்டுகளில் நுண்ணிழைகள் ஆக்டின் இழைகள், கோல்கை உறுப்புகளிலிருந்து தோன்றும் வெசிக்கிள்கள், எண்டோபிளாச வலை ஆகியவை காணப்படுகிறது. கார்போஹைட்ரேட்டுகளான பெக்டின் மற்றும் ஹெமிசெல்லுலோஸ்சைப் பெற்றுப் பிராக்மோபிளா ஸ்டின் நுண்ணிழைகளோடு நகர்ந்து மையப்பகுதியில் இணைந்து புதிய பிளாஸ்மா சவ்வினை உருவாக்குகிறது. செல் சுவர் உருவாக்கத்தில் முதல்நிலையானது, புதிதாகத் தோன்றிய செல்களுக்கு நடுவில் ஒரு கோடு உண்டாகின்றது. இதற்குச் செல் தட்டு என்று பெயர். செல்லிற்குள் செல் தட்டு விரிவடைந்து மையத்தட்டு உருவாகிறது. மையத்தட்டின் இருபுறமும் செல்லுலோசினால் ஆன புதிய செல் சுவர்களை இரு தாவரச் செல்களுக்கிடையே உருவாகிறது.
குன்றல் பகுப்பு (மியாசிஸ்):
மீயோம் என்ற கிரேக்கச் சொல்லிற்குக் குன்றல் என்று பொருள்படும். எனவே இது குன்றல் பகுப்பு எனப்படுகிறது. இப்பகுப்பில் குரோமோசோம்க ள் இணைசேரும் நிகழ்வான சினாப்சிஸ் காணப்படுவது இதன் சிறப்பாகும். அத்துடன் குரோமோசோம்களின் எண்ணிக்கையில் குறைதலும் குறிப்பிடத்தக்கது. பாலினப் பெருக்கத்தில் பங்காற்றும் உறுப்புகளின் திசுக்களில் இப்பகுப்பு நிகழ்கிறது. இதன் விளைவாக உருவாக்க ப்படும் கேமீட்டுகளில் தாய் செல்லின் குரோமோசோம்களின் எண்ணிக்கையில் பாதியளவாகக் குரோமோசோம்க ளின் எண்ணிக்கை காணப்படும். எனவே புதிய மரபுச் சேர்க்கை அடைந்த வகைகளை உருவாக்குவதில் இப்பகுப்பு முக்கியப் பங்காற்றுகிறது.
விலங்குகளில் விந்தகத்தில் ஹாப்லாய்டு விந்துக்கள் உருவாக்கவும் அண்டகத்தில் ஹாப்லாய்டு முட்டைகள் உருவாக்கவும் இப்பகுப்பு உதவுகிறது.
பூக்கும் தாவரங்களில் மகரந்தப் பைகளில் நிகழும் மைக்ரோஸ்போர் ஆக்கத்தின் போது, சூலில் நிகழும் மெகாஸ்போர் ஆக்கதின் போதும் இப்பகுப்பு நிகழ்கிறது. மைட்டா சிஸ் பகுப்பைப் போல் இல்லாமல், இப்பகுப்பில் மரபியலில் ஒவ்வாத சேய் செல்கள் உருவாக்கப்படுகின்றன. எனவே புதிய மரபுச் சேர்க்கை அடைந்த வகைகளை உருவாக்க இப்பகுப்பு முக்கியப் பங்காற்றுகிறது.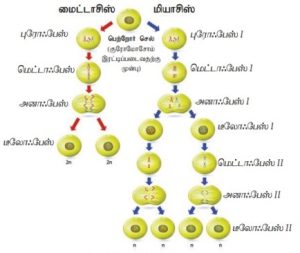
ஒடுக்கற்பிரிவு பகுப்பின் நிலைகள்:
ஒடுக்கற்பிரிவை இரண்டு பிரிவுகளின் கீழ் ஆய்வு செய்யலாம், அதாவது ஒடுக்கற்பிரிவு I மற்றும் ஒடுக்கற்பிரிவு II. மைட்டோசிஸைப் போலவே, செல் பிளவுபடாதபோது இடைநிலையில் இருப்பதாகக் கூறப்படுகிறது.
ஒடுக்கற்பிரிவு I – குன்றல் பகுப்பு:
புரோஃபேஸ் I: நீண்ட கால அளவு கொண்டுள்ளது. இது ஐந்து துணை நிலைகளாகப் பிரிக்கப்பட்டுள்ளது. அவையாவன: லெப்டோட்டீன், சைக்கோட்டீன், பாக்கிடீன், டிப்ளோட்டீன், டையாகைனசிஸ்.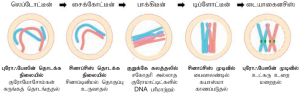
- லெப்டோட்டீன்: இந்தத் துணை நிலையில் குரோமோசோம்கள் ஒளி நுண்ணோக்கி மூலம் எளிதில் காணக்கூடியதாக உள்ளன. குரோமோசோம்கள் சுருங்கிக்குறுகுதல் நிகழ்கிறது. சகோதரி குரோமாட்டிட்கள் சுருங்குவதே இதற்குக் காரணமாகும்.
- சைக்கோட்டீன்: ஒத்திசைவு குரோமோசோம்கள் இத்துணை நிலையில் இணைசேர்கின்றன. இதற்குச் சினாப்சிஸ் என்று பெயர். இந்த சினாப் சிஸ் நிகழ்வு சினாப்டினிமல் தொகுப்பின் (Synaptonemal complex) உதவியால் ஏற்படுகிறது. இதனால் தோன்றும் இணை குரோமோசோம்களின் தொகுப்பிற்குப்பைவாலண்ட் என்று பெயர். இதில் இரு குரோமோசோம்களின் நான்கு குரோமாட்டிட்கள் தொகுதியடைவதால் இது நான்கமை நிலை (Tetrads) எனப்படுகிறது.
- பாக்கிடீன்: இந்த நிலையில் பைவாலண்ட் குரோமோசோம்களின் நான்கமை நிலை (Tetrads) தெளிவாகப் புலப்படுகிறது. மியாசிஸ் – Iல் பைவாலண்ட் ஒவ்வொன்றும் 4 குரோமாட்டிட்கள், 2 சென்ட்ரோமியர்களைக் கொண்டுள்ளது. ஒத்திசைவு குரோமோசோமின் சகோதரி குரோமாட்டிட்கள் குறுக்கெதிர் மாற்றம் (Crossing over) நடைப்பெற்ற பகுதியில் மீள்சேர்க்கைக்கு உதவும் இலக்குகள் (Recombination nodules) தோன்றுகின்றன. இந்தத் துணை நிலையின் முடிவில் ஒத்திசைவு குரோமோசோம்களுக்கிடையே மீள் சேர்க்கை நிகழ்வது முடிவுற்றுக் குறுக்கெதிர் மாற்றம் நடந்தபகுதியில் மட்டும் குரோமோசோம்கள் இணைந்துள்ள நிலை ஏற்படுகிறது. இந்நிகழ்விற்கு ரிகாம்பினேஸ் என்ற நொதி உதவுகிறது.
- டிப்லோட்டீன்: சினாப்டினிமல் தொகுப்பு கலைந்து கரையத் தொடங்குகிறது. குறுக்கெதிர் மாற்றம் நடந்து, ஒன்று அல்லது அதற்கு மேற்பட்ட புள்ளிகளில் ஒத்திசைவு குரோமோசோம்கள் பிணைந்த நிலையிலேயே உள்ளன. இவ்விலக்கில் “X” வடிவ அமைப்பு காணப்படுகிறது. இவ்விலக்குகள் கயாஸ்மாக்கள் எனப்படுகின்றன. குரோமோசோம்களில் மீள் சேர்க்கை நிகழ்ந்த இலக்கை இந்தக் கயாஸ்மாக்கள் குறிக்கின்றன. சகோதரி குரோமாட்டிட்கள் நெருக்கமாக இணைவுற்றிருந்தாலும் ஒத்திசைவு குரோமோசோம்கள் ஒன்றை விட்டு ஒன்று விலகிய நிலையில் காணப்படும். இருப்பினும் இவை கயாஸ்மா இலக்குகளில் இணைந்தே காணப்படுகின்றன. இந்த துணை நிலையில் பால் தன்மை மற்றும் உயிரிகளுக்கேற்ப நாட்கள் அல்லது வருடங்கள் வரை நீடிக்கும்.
- டயாகைனசிஸ்: காயஸ்மாக்கள் முடிவுறுதல் இத்துணை நிலையில் நிகழ்கிறது. கதிர்கோல் இழைகள் கூடுகின்றன. உட்கரு உறை சிதையத் தொடங்குகிறது. ஒத்திசைவு குரோமோசோம்கள் குறுகிச் செறிவடைகின்றன. நியுக்ளியோலஸ் மறைகிறது.
மெட்டாஃபேஸ் I:
இரண்டு ஒத்திசை குரோமோசோம்களின் சென்ட்ரோமியருடன் கதிர்கோல் இழைகள் இணைகின்றன. இணை சேர்ந்த பைவாலண்டுகள் செல்லின் மையப் பகுதியில் அமைகின்றன. இதற்கு மெட்டாஃபேஸ் தட்டு என்று பெயர். மெட்டாஃபேஸ் தட்டில் உள்ள ஒத்திசைவு குரோமோசோம்கள், சீரற்ற பரவல் காரணமாகச் சார்பின்றி ஒதுங்குதல் நடைபெறுகிறது.
அனாஃபேஸ் I:
ஒத்திசைவு குரோமோசோம்கள் ஒன்றை விட்டு ஒன்று பிரிதல் இந்நிலையில் நிகழ்கிறது. கதிர்கோல் இழைகள் சுருங்குவதால் இது ஏற்படுகிறது. ஒவ்வொரு ஒத்திசைவு குரோமோசோம் இணைகளில் உள்ள இரண்டு குரோமாட் டிட்களும் பகுபடாத முழுச் சென்ட்ரோமியரும் செல்லில் எதிரெதிர் துருவங்களை நோக்கிச் சென்றடைகின்றன. குரோமோசோம்களின் எண்ணிக்கை சரிபாதியாகக் குறைவது இந்நிலையில் தான் நிகழ்கிறது. எதிரெதிர் துருவங்களை அடைந்த ஒத்த குரோமோசோமில் ஒன்று தாய்வழி வந்ததாகவோ அல்லது தந்தை வழி வந்ததாகவோ இருக்கிறது. சகோதரி குரோமாட்டிட்கள் சென்ட்ரோமியருடன் இணைந்து காணப்படுவது குறிப்பிடத்தக்கது.
டீலோஃபேஸ் I:
ஒவ்வொரு துருவத்திலும் ஹாப்லாய்டு குரோமோசோம் தொகுப்பு காணப்படுகிறது. இதனால் ஹாப்லாய்டு எண்ணிக்கையுடைய இரு சேய் செல்கள் உருவாக முடிகிறது, துருவத்திலுள்ள ஒவ்வொரு தொகுப்பையும் சவ்வு சூழ்வதால் ஒரு உட்கரு உருவாகிறது. இவ்வுட்கருவில் குரோமோசோம்கள் குரோமாட்டின் இழைகளாக மாறுவதுடன் நியூக்ளியோலசும் உருவாகிறது.
தாவரங்களில் குன்றல் பகுப்பின் போது காரியோகைனசிஸ் அடுத்து சைட்டோகைனசிஸ் நிகழ்வதால் செல்தட்டு உண்டாகி, இரு சேய்செல்கள் உருவாகின்றன. இந்நிலைக்கு இரு செல் நிலை என்று பெயர். இரண்டு மியாசிஸ் பகுப்பிற்கும் இடையே குறுகிய காலத்தில் அமைந்த ஒரு நிலை உருவாகிறது. இதற்குப் பகுப்பிடைக்காலம் என்று பெயர்.
மியாசிஸ் II – சமநிலை பகுப்பு:
இந்த பிரிவு மைட்டோடிக் ஒடுக்கற்பிரிவு என்று அழைக்கப்படுகிறது, ஏனெனில் இது மைட்டோசிஸை ஒத்திருக்கிறது. இது மைட்டோடிக் பிரிவுகளின் அனைத்து நிலைகளையும் உள்ளடக்கியதால்.
புரோஃபேஸ் II:
இரண்டு குரோமாட்டிட்களை கொண்ட குரோமோசோம் குட்டையாகி, சுருங்கி, அடர்த்தி அடைந்து, கண்ணுக்குப் புலப்படக்கூடியதாக உள்ளன. உட்கரு சவ்வு மற்றும் நியூக்ளியோலஸ் மறைகின்றன. இதனைத் தொடர்ந்து புதிய கதிர்கோல் இழைகள் செல்லின் அச்சிற்குக் குறுக்காக அமைந்த இரு துருவங்களிலிருந்து தோன்றுகின்றன.
மெட்டாஃபேஸ் II:
ஒவ்வொரு செல்லிலும் உள்ள ஒத்திசைவற்ற குரோமோசோம்கள் கதிர்கோல் இழைகளுக்குக் குறுக்கே அமைந்த மையத்தட்டில் அமைந்து மெட்டாஃபேஸ் தட்டு ஒன்று தோன்றுகிறது. கதிர்கோல் இழைகள் சகோதரி குரோமாட்டிட்களின் சென்ட்ரோமியருடன் பிணைகின்றன.
அனாஃபேஸ் II:
ஒவ்வொரு குரோமோசோமின் சென்ட்ரோமியரும் துண்டிக்கப்படுவதால் அதன் சகோதரி குரோமாட்டிட்கள் பிரிந்து துருவங்களை நோக்கி நகர்கின்றன. இது கதிர்கோல் இழைகள் சுருங்குவதால் நிகழ்கிறது.
டீலோஃபேஸ் II:
இந்நிலையில் ஹாப்லாய்டு குரோமோசோம்களைப் பெற்ற நான்கு உட்கரு உருவாகின்றன. கதிர்கோல் இழைகள் மறைகின்றன. உட்கரு உறை மற்றும் நியூக்ளியோலஸ் மீண்டும் உருவாகிறது. இந்த உட்கரு பகுப்பு முடிவுற்றதும் சைட்டோபிளாச பகுப்பு நிகழத் தொடங்குகிறது. செல்தட்டுகள் தோன்றி நான்கு ஒற்றை மடங்கு குரோமோசோம்களை கொண்ட சேய் செல்கள் உருவாகின்றன. இதற்கு நான்கு செல் நிலை என்று பெயர்.
ஒடுக்கற்பிரிவின் முக்கியத்துவம்:
இது உயிரினங்களில் நிலையான எண்ணிக்கையிலான குரோமோசோம்களை பராமரிக்கிறது.
கிராசிங் ஓவர் நடைபெறுகிறது மற்றும் மரபணு பொருட்களின் பரிமாற்றம் இனங்கள் மத்தியில் மாறுபாடுகளுக்கு வழிவகுக்கிறது. இந்த மாறுபாடுகள் பரிணாம வளர்ச்சிக்கான மூலப்பொருளாகும். ஒடுக்கற்பிரிவு மரபணு மாறுபாட்டிற்கு வழிவகுக்கிறது, இது மரபணுக்களின் வெவ்வேறு சேர்க்கைகளை சுயாதீன வகைப்படுத்தலின் மூலம் கேமட்களாகப் பிரிக்கிறது.
பல்வேறு சுற்றுச்சூழல் அழுத்தங்களுக்கு உயிரினங்களின் தழுவல்.
பாதி – தற்சார்பு:
அரை தன்னாட்சி உறுப்புகள் என்பது அவற்றின் சொந்த DNAவைக் கொண்டிருக்கும் உறுப்புகளாகும் மற்றும் கருவில் இருந்து சுயாதீனமாக இனப்பெருக்கம் செய்கின்றன. குளோரோபிளாஸ்ட் மற்றும் மைட்டோகாண்ட்ரியா அரை தன்னாட்சி உறுப்புகள் என்று அழைக்கப்படுகின்றன, ஏனெனில் அவை அவற்றின் சொந்த மரபணு பொருள் (DNA) மற்றும் அவற்றின் செயல்பாட்டிற்கு தேவையான புரதங்களை ஒருங்கிணைக்கும் திறன் கொண்டவை.